Chapter Nine
CHARACTERISTICS
The founding of agricultural practices on ecological principles, whether deliberate or not, is certainly not new; two thousand years ago the Romans had considered ecological factors in agricultural management, and groups of swidden farmers in remote areas today farm in a sustainable and environmentally benign manner. Agricultural development is marked by periods, however, when linkages between components of agroecosystems (agricultural ecosystems) and the fragile ecological processes have been broken. Indonesian examples of the recent past would include the use of increasing quantities of the biocide1 DDT during the 1970s to control increasingly resistant bagworms (larvae of psychid moths) in oil palm plantations, the planning of some transmigration settlements in totally unsuitable locations, and the adoption of inappropriate farm models. Inattention to ecology has resulted in recurrent outbreaks of pests and diseases (p. 293), soil erosion, declining soil quality, and pollution. It is now realized that these problems cannot be dealt with one by one, for they are linked by ecological, social and economic processes. For this reason systematic ecological analyses of agroecosystems are a new and effective means of research and development (Conway 1985a, b).
Agroecosystems are in some sense simpler than natural ecosystems because there is limited influence from other systems, the boundaries can be defined, and the numbers of species present and physical processes are fewer, but the large number of management and control decisions (both correct and incorrect) by the farmer introduces great complexity.
Any ecosystem can be described in terms of certain measurable characteristics, and those of most concern to agroecosystems are productivity, stability and resilience (or sustainability). Productivity is the net gain of a sought item over time and, in the agricultural sense, can be measured as annual yield of a crop or net income. Stability is a measure of the variation of production caused by normal, small-scale variations in environmental or economic conditions. Resilience, or sustainability, is the ability of a system to maintain or regain its productivity after a period of stress or a sudden major environmental or economic change. These three can be assessed for any ecosystem, but a fourth characteristic, equitability, is really only of relevance to agroecosystems. Equitability is the evenness with which the products, or the income derived from it, of the agroecosystem are distributed among the members of the farm, village, region or nation (fig. 9.1) (Conway 1985a, b).
Different agroecosystems can be described in terms of the above four parameters. Agricultural developments tend to be aimed at increasing productivity but, as can be seen, this is achieved at the expense of equitability and sustainability. Thus the different agroecosystems examined briefly in this chapter have different ecological properties (table 9.1).


Figure 9.1. Performance of agroecosystems under conditions of high and low productivity, stability, sustainability and equitability.
After Conway 1985a, b
SWIDDEN, SHIFTING AND INTENSIVE AGRICULTURE
Swidden agriculture,2 in its purest sense, is the repeated use of a patch of forested land for the cultivation of crops, and is characterized by long fallow periods between short periods of intensive production. One of the few groups of Sulawesi farmers which still practices traditional swidden cultivation is the Laoceh people who live on the slopes of Mt. Ogoamas, Donggala. Swidden agriculture is an extensive form of shifting agriculture and is not the same, despite what one might be led to believe, as the generally unsustainable slash and burn or intensive shifting cultivation practised in many areas (fig. 9.2). Swidden agriculture proceeds as follows: an area of forest is cleared, the remains are left to dry and these are then burned. Crops are subsequently sown in the ashes between the remaining large tree trunks. Some nutrients are lost in the smoke from the fire, pH is lowered and rain washes much of the ash into rivers before it has had a chance to become incorporated into the soil (Geertz 1963). Other nutrients are made available to plants by burning the organic matter (Andriesse and Koopmans 1984). What erosion does take place (due to the mosaic pattern of surrounding fallowed land), is usually trapped by vegetation at the field boundary (Hamilton and King 1983). After one or two crops, yields decrease, weeds become a serious problem (Chapman 1975) and the area is abandoned for 15-30 years during which time the soil recovers (Soeri-anegara 1970; van Baren 1975). The farmer then moves to an area of building-phase forest (p. 359). This is generally the site of a former field, because this type of forest has a smaller stature and is easier to clear than primary forest. This old field may contain mature fruit trees, such as durian, the fruit of which would have been harvested in the intervening years.
The use of cleared and burned land for swidden agriculture alters the early stages of forest succession (p. 359); weeding removes shoots growing from tree stumps and although efforts are made to remove herbaceous plants, their numbers increase. These herbs dominate the area soon after the land is abandoned, but fast-growing pioneer trees begin to smother the smaller plants after a year (Symington 1933; Uhl et al. 1982). Herbs and pioneer trees dominate these cultivated areas because they have many viable seeds in the soil and/or the seeds are easily dispersed (p. 362). It is clear that when the swidden agriculture cycle is long enough (e.g., 30 years), species of herbaceous weeds are not present in the secondary forest felled prior to cultivation. When the cycle is reduced to, say, just five years and succession has not been allowed to progress very far, domination by herbaceous weeds during even the first crop can become a serious problem.
Swidden agriculture, of the type described above, can support 30-40 individuals/km2 and depends for its success on the farmers harvesting for subsistence rather than for cash crops and on cultural restraints that have evolved as part of the traditions bound up in animistic religions. It is possible, however, to have a rather over-romantic idea of swidden agriculture and there is a mistaken belief that all people distant from missionary or government influence are good stewards of their environment. For example, in land occupied by the Kayu Merangka3 group of the tribal Towana people in the remoter parts of the Morowali region of Central Sulawesi, there are vast areas of grassland caused by shifting cultivation on poor soils (Powell n.d.). Similarly, the Pipi Koro region west of the headwaters of the Palu River, also in Central Sulawesi, is desolate grassland, but in and around the nearby Lore Lindu National Park, where settled, rice-based agriculture was introduced at the start of the century, large areas of magnificent forest remain (Anon. 1981). It is not known what 'went wrong' in these areas but it may be that the farmers originally worked land in the richer lowlands but were forced by various pressures to to move to the poorer soils of the hills where their techniques were inappropriate.

Figure 9.2. The distinction between traditional swidden agriculture (a), and the now prevalent shifting agriculture (b) which is endangering so many forested areas.
After Rijksen 1978
With increasing populations and closer contact with the outside world, the restraints of swidden agriculturalists are eroded (Geertz 1963), and it is clear that the environmental care implicit in Christianity and Islam (Schaeffer 1970; Sutopo and Suayeb 1980) does not seem to find its way into the lives of new generations of converted shifting cultivators. Indeed, traditional, extensive swidden agriculture in its true sense has virtually disappeared from Sulawesi and a very small percentage of the population is involved. Published figures for areas under swidden or shifting agriculture do not as a rule include definitions of the terms used or differentiate between the two systems and are therefore of little use.
Shifting agriculture, as a term, thus covers a wide range of farming practices causing varying degrees of damage to the environment but, in general, it is practised with almost no cultural or social restraints and the cultivation of cash rather than subsistence crops is the major use of cleared land. Rested land is scarce and mature-phase forest is usually cleared when the former cultivated area is exhausted or when alang-alang grass can be kept at bay no longer by simple (cheap) weeding. Regeneration to mature-phase forest in such areas would take many centuries (p. 360).
Irrigated rice culture, introduced to Indonesia from India about 1,000 years ago is clearly regarded at most levels of society as the ultimate, the most rational, and most advanced form of agriculture for Indonesia. While recognizing the major role that rice plays in feeding the people of Indonesia, it is instructive, however, to stand back and to examine the applicability of rice growing for all people in all places. The agricultural system seen as the opposite end of the spectrum of modernity, is swidden agriculture, yet when this, and even some shifting agriculture, is subject to detailed analysis, they appear to be rational uses of manpower and land. For example, a typical Indonesian upland rice field may yield more rice per man-day than a typical wet rice field. Thus the rejection by such hill swidden farmers of incentives and invitations to change their agricultural system may be based on very sound economic motives, not just on backward thinking or fear of change.
Detailed study has shown how some swidden farmers are rational and sophisticated users of their natural environment (Dove 1985a). This is important to realize, since most of the blithe, blanket statements concerning swidden and shifting agriculture indicate that it is based on ignorance and inevitably results in misuse of the environment. It is this view that supports resettlement programs of remote groups of people (Satjapradja and Mas'ud 1978; Mile and Semadi 1981), but there is no compelling environmental reason to move swidden agriculturalists if they are engaged in stable, sustainable farming (Hamilton and King 1983). Among one group of Towana practising swidden/shifting agriculture between Luwuk and Ampana, there are former kampong leaders, former Muslims and Christians, and men who had been educated in Dutch schools. These people know about the methods of intensive agriculture but have decided to keep their options open and maintain an agricultural system whose benefits and advantages in their hilly environment they know (Atkinson 1979; Hidayan 1980).
The intensive nature of rice farming, promoted since the days of the early Javanese kingdoms, actually makes poor use of manpower and limits farmers' opportunities for planting cash crops. It is interesting, for example, that in the authoritative book on Indonesian rice production (Mears 1981) there are abundant data on yields per hectare, but not a single set of data on yields per man day. High yields per unit area are clearly favourable in areas of high population density, where labour is abundant and suitable land is scarce, but they are less important where those conditions do not apply. It is argued that swidden agriculture is incapable of absorbing continual increases in population density, but the production of rice from irrigated rice fields tends to decline as the population density increases (Geertz 1963). Absorption of manpower is not necessarily a superior goal or a better answer to demographic and economic problems than productivity of manpower (Dove 1985b).
So much could be done to breed improved strains of crops grown by shifting agriculturalists, to introduce new species, and to discover the best species for reforestation of fallow slopes. Agricultural intensification is neither a natural nor an inevitable development even in areas where environmental conditions make it possible. For example, alluvial soils in many Sulawesi valleys and along many coastal strips were not used for intensive agriculture until this was effectively imposed by the colonial and national governments. Conversely, agricultural intensification can and does occur as a result of government programs in areas whose physical, biological and social environments are not suitable. For example, the large Wawotobi dam being built near Una-aha, Kendari, is intended to allow the irrigation of 18,000 ha of existing and future rice fields. The Tolaki, the dominant group of people in the Kendari area, are dry-land farmers or shifting agriculturalists and have shown little inclination in the past to change to wetland rice culture. In addition, other irrigation schemes in Kendari County have foundered because the actual water availability was not as great as was predicted. This experience is shared by certain irrigation schemes in seasonal parts of Gorontalo (Anon. 1986).
PEST ECOLOGY AND CONTROL
It is generally true that in their natural ecosystem, species that become agricultural pests are present at relatively low densities, tend to have specialized and ephemeral niches, and have populations that never reach pest proportions. With a change in land use, however, a niche that was once restricted can become available continuously over wide areas, and natural predators of the pest may not be able to hold the growing population in check if suitable habitat or other vital resources are not available. An insect predator may be dependent, for example, on certain plants on which its larvae have to feed.
The spraying of biocides onto crops is the conventional modern way of controlling pests.4 Continued use can lead to resistance as evidenced by the hundreds of insects, mites and ticks that have developed resistance to one of more biocides. The resistance spreads through populations because the individuals that survive the effects of the biocide by behavioural, biochemical or physiological means, pass on their genetic adaptations to the next generation. Repeated applications of a biocide may thus lead to resistant populations. There is a greater tendency for species with a high intrinsic rate of population increase (the r-strategists-p. 345) to develop resistance, but the main determining factor is frequent application of high concentrations of biocides. Once a species has developed resistance to a biocide—and resistance has been documented for all chemical groups— the state is more or less irreversible. It is therefore quite conceivable that resistance to all known biocide groups may one day be found in populations of certain insect strains (Conway 1982). Resistance does not pose an immediate threat to world food production but it can cause extremely serious problems on a local level. In one of the latest reviews of biocide resistance, Asia is identified as the area most likely to suffer crises of pest resistance, particularly if the use of biocides grows to the high levels currently used in countries such as Japan (Conway 1982).
During the 1960s and early 1970s the potential dangers and undesirable effects of biocides were brought to the notice of the general public. Much was overstated or biased and it was impractically suggested that controlling pests by biological means was the only acceptable path to follow. Biological control is a term used for pest control by predators, parasites, pathogens, behaviour-changing hormones, sterilization of male pests, or by using resistant crop varieties. Most biological controls are highly selective and do not cause environmental damage. Other advantages are that a controlling organism can seek out its prey in a way that chemicals cannot, can increase in number and spread, and resistance to the predator by the prey is either slow or unlikely to develop. Disadvantages are that control is slow, pests are almost never wholly exterminated, patterns of control are often unpredictable, techniques are expensive to develop properly and to apply, and expert supervision is required. In addition, problems can occur when pathogens are imported along with a controlling predator or parasite or when the agent is less specific than was hoped. These disadvantages were not emphasized during the early chemical scares and society was presented with an unrealistic choice between biocides on the one side and apparently perfect biological control on the other. Fortunately, agriculturalists did not make such simplistic choices and instead have developed the concept of integrated pest control.

Figure 9.3. Increase in the use of biocides (000s tonnes) in South Sulawesi between 1970 and 1984.
From Anon. 1982
Integrated pest control was defined over 25 years ago as "applied pest control which combines and integrates biological and chemical control. Chemical control is used as necessary and in a manner which is least disruptive to biological control" (Stern et al. 1959), and despite much research it does not receive the committed support it deserves (Conway 1985a). This is reflected in the huge increase in the use of biocides (fig. 9.3). One problem in pest management was that until recently only very few types of biocides were available and these were incapable of controlling the full range of insect pests. Improper use has led to an unimpressive impact and a consequent lack of enthusiasm for biocides on the part of the farmers. Certain pest problems or certain stages of those problems can only really be overcome using chemicals. Other methods can serve to keep most pest populations within limits for most of the time. These methods include making the agricultural environment as favourable as possible to predators and as unfavourable as possible to pests, and the various forms of biological control described above. When pest populations reach a certain predetermined threshold level judged capable of precipitating a serious economic loss, however, selective chemicals need to be used (p. 599).
It is notable that accounts of pests and the problems they cause often close with the remark that if the ecology of pests predators and crops (or related wild relatives) (Salick 1983) were better understood, better control measures could be developed. In theory, if the pest and its position in a food web is thoroughly understood, then accurate predictions can be made concerning the probable effects of different forms of control. The gathering of data which could genuinely achieve such ends for just a single pest species could take many years (van Emden 1974; Heckman 1979), during which time serious increases in population levels could occur. Thorough documentation and analysis of the different ecological responses of pest populations to different forms of control should lead to better selection of control programmes and the effects of such programs could be predicted with increasing accuracy (Altieri et al. 1983). In this way, ecological knowledge of the organisms concerned will lead to effective and environmentally sound pest control.
Predatory spiders have been all but forgotten in discussions on biological control, because little is known of their ecology and so they have not received the attention they deserve. Their main characteristic is that they are generalist rather than specialist predators but agricultural entomologists have rather sought the parasite or predator to control the pest under scrutinity. Evidence suggests that spiders can reduce pest populations significantly in rice and cotton crops, but it is the community of spiders which needs to be encouraged, not just a single species. Safeguarding the spiders can be achieved simply by paying attention to their habits; for example, spraying at mid day when spiders tend to be inactive and under some form of shelter (Riechert and Lockley 1984).
Most of this section concerns the control of insect pests, but rats and birds are ubiquitous pests which also cause considerable damage (p. 583). Rats are considered to be the most important group of pests hindering agricultural production in Southeast Asia (Soerjani 1980). Rats are known to occur in almost all agricultural crops and to damage many stored products (Anon. 1976, 1980; Myllymaki 1979; Estioko 1980; Soekarna, et al. 1980). They present the additional hazard of being a health risk to humans; the organisms causing scrub typhus, meningoencephalitis, plague and lepto-spirosis are all known to be carried by parasites of agricultural or urban rats (Lim et al. 1980; Sustriayu 1980). For example, the mite Leptotrombidium deliense known as a vector of scrub typhus has been found on rats caught in Central Sulawesi and Bogani Nani Wartabone National Park (van Peenan et al. 1974; Durden 1986). Luckily, in this and many other cases, even though the disease and the organisms may be found close to where humans work or live, the disease does not necessarily break out. The environmental conditions required for the disease to be transmitted are not known, but it is clear that detailed ecological knowledge of rats should be collected now, rather than once an epidemic has begun.
The number of bird species inhabiting rice fields, plantations and other areas of permanent agriculture is quite considerable but none of them is an inhabitant of mature- or building-phase lowland forests. This serves to demonstrate how few of the forest birds can survive in disturbed areas (p. 434).
RICE FIELDS
Introduction
Much of this section is concerned with, and uses data from, South Sulawesi. This is because South Sulawesi is a major rice-growing area with over three times as much land under rice as all the other Sulawesi provinces combined. Indeed, South Sulawesi is one of only two Indonesian provinces that produces a yearly rice surplus. Just over half its total arable land (about 1,100,000 ha) is committed to rice, and the next most planted crop is maize which covers just 13% of the rice area. About 50% of the rice is rain-fed, 45% is irrigated by various means, and just 5% is unirrigated in upland areas. Of the rain-fed and irrigated rice5 most is grown in the alluvial coastal plains which have the most suitable soils but even these may be deficient in nitrogen and sulphur (van Halteren 1979).
In some karst areas of Indonesia, such as the famous Mt. Sewu area of the Special Region of Yogyakarta, rice culture is more or less impossible because of the porous nature of the bedrock. In the Bone cockpit karst area (p. 470), the topography is superficially similar to that of Mt. Sewu but not far below the surface are impermeable rocks which can cause ponds to form on the surface. In this area very simple irrigation can produce two crops of rice per year (fig. 9.4). The hill slopes are used for tobacco and after just one year under this crop, the fields are left fallow for three to five years and so there is less soil exposure and hence less erosion. The human population densities of Mt. Sewu and the Bone karst area are more or less the same (about 330 people/km2) but the environmental conditions in Bone are far more favourable thanks to the underlying rock (Uhlig 1980).

Figure 9.4. Bone and Maros karst regions showing their agricultural patterns, a -favourable agricultural area with karst springs (irrigation, two rice harvests, or one rice and one tobacco harvest, per year); b - rice fields with technical irrigation (one rice harvest per year); c - rice fields with impounded rainfall or simple ditch irrigation (one rice harvest, during the rainy season); d - terraces with permanent dry-field cultivation; e - shifting cultivation; f - pioneer-phase forest, grassland; g - rice fields with impounded or simple ditch irrigation (one rice harvest, during the rainy season).
After Uhlig 1980
The plain around the tower karst of Maros is densely settled and at first sight the settlement and cultivation of the area is very similar to most other rice growing areas of South Sulawesi. The main difference though, is the karst springs at the base of the cliffs which provide a year-round supply of water and two crops per year can be grown using simple irrigation ditches. Water from the relatively large Bantimurung River issuing from the karst of Maros also has a reliable year-round flow and has been routed to provide technical irrigation to 70 km2 of rice fields—one of the largest irrigation schemes in Sulawesi (Uhlig 1980).
Rice Fields as an Ecosystem
Rice fields are essentially modified swamps and the animals and plants they contain are typical of such ecosystems. Rice fields as an ecosystem are maintained by humans orchestrating the yearly or twice-yearly cycles of flooding, ploughing, removal of organic matter (harvesting) and enrichment with fertilizers. If people were to relax their efforts the condition of the rice field would very rapidly change and enter the forest growth cycle (p. 359). A rice field is generally by no means a monoculture. Apart from the rice crop, people also harvest kangkung Ipomoea aquatica (p. 269) as a vegetable, snails, prawns, crabs, fish and frogs. In certain areas, such as Tana Toraja, the centre of rice fields are deliberately deepened so that, after harvesting when the field is allowed to dry out, a pond is formed. These ponds are a refuge for aquatic organisms and a source of food for ducks.
Rice fields as an ecological system have hardly been studied at all and work has concentrated instead on the important subject of pest control and fish culture. Fish culture could most easily be enhanced, however, by increasing the productivity of the aquatic organisms on which they feed and this is only possible through an understanding of how rice fields function in an ecological sense.
Comprehensive analyses of the fauna6 and flora have been conducted, however, in Peninsular Malaysia (Fernando 1977) and northeast Thailand (Heckman 1979). The site chosen for the detailed Thai study was near Udorn Thani at about 17°N which has a very seasonal climate with three months when virtually no rain falls and minimum temperatures in the dry months fall to 5°C. Consequently, many species common in more stable environments in Thailand and elsewhere in Southeast Asia are absent or rare in this study area, but the rice fields are by no means depauperate in species. One reason for this may be the fact that rice had probably been grown in the area for at least 5,500 years. In areas where rice has been grown for only a 100 years or so the number of animals and plants is somewhat low but, with time, adaptations by other species are likely to occur and the species total will increase. Some animals, such as certain fish, are deliberately introduced into the ecosystem and a number of these have been cultured for food for at least 2,000 years.
The study in Thailand found that a single field was inhabited by a staggering 589 species of plants and animals, 209 of which were considered rare. This species total included 38 flowering plants, 173 algae, 120 protozoans, 52 rotifers, 12 molluscs, 33 crustaceans, 21 dragonflies, 39 beetles and 18 fish. No real plankton are present, possibly because the water is simply too shallow and too full of plant material to allow such species to survive. Mosquitoes are rare in rice fields where the water is more than a few centimetres deep because predatory fish will soon consume them (Heckman 1979). The manner in which these organisms divide up the available resources is scarcely understood although it is easy enough with fishes (table 9.2).
Most of the larger aquatic animals are adapted to living in water with little or no dissolved oxygen. The fish have accessory breathing organs, the snails have lungs and gills, some snails live in close contact with the oxygen-rich surface layer, some insects and spiders carry a bubble of air with them below the surface, while others may store oxygen in their tissues.
The periodic drying of the fields means that newly-wet fields have to be recolonized or that plants or animals need resistant, dormant stages to endure the 'drought'. Some species such as snails, crabs, the swamp eel Monopterus alba and even an Enhydris snake (p. 305) bury themselves in the mud and remain inactive during the dry period, some move to permanent water bodies and many die. Those that die may leave resistant eggs, spores or seeds on the mud. Recolonisation can occur from nearby swamps, rivers or lakes, but in areas where virtually all swamps have been converted, the ricefield fauna is relatively poor (Fernando et al. 1980).
Most agronomists are charged with increasing the yield of rice per unit area. Where this involves the use of chemicals in pest control, insufficient attention is usually given to the other productive components of the rice fields. Great care must be taken in the application of chemicals so as not to distort or break the food web in a way that would reduce the capacity of a rice field to produce protein. A few organisms may even be encouraged to live alongside the rice: an example is the small fern Azolla pinnata (Azol.) which has clearly useful nitrogen-fixing blue-green algae Anabaena azollae in its leaves and is also used as animal feed. This inattention is regrettable since low species richness and high population densities of few species are often regarded as symptoms of an ecosystem under stress. In Peninsular Malaysia improved rice yields achieved through the use of biocides and double-cropping have been shown to be associated with reduction of the associated aquatic fauna (Yunus and Lim 1971; Fernando 1977).

After Heckman 1979
This may seem defensible where there are pressures to over produce and to export but, for many farmers subsistence is the major goal. Chemicals may result in high yields, but if the farmers are unable to benefit from what can be, and perhaps should be, a multi-culture agroecosystem, effort and time must be spent producing alternatives to the varied protein and vitamin sources of a diverse and healthy rice field.
Insect Pests on Rice
All parts of a rice plant (except roots under water) at all stages of growth are attacked by insects (fig. 9.5) and the roots are attacked by nematode worms. Some, such as grasshoppers, feed on rice and many other plants while others, such as certain leafhoppers are known only from rice. The insect pests on rice in South Sulawesi have been studied in detail and the white stem borer Tryporyza innotata is the most important followed by the rice seedbug Leptocoria oratoria and the rice leaf folder Cnaphalocrosis medinalis. Other important insect pests include the striped stem borer Chilo suppresalis, the green leafhopper Nephotettix virescens, and the brown planthopper Nilapawata lugens. It is not the actual damage done by the leafhoppers to the plants that causes concern, but rather the virus diseases they spread. The green leafhopper transmits the tungro virus which devastated large areas of rice around Palu and in South Sulawesi from 1972 to 1974. The brown planthopper transmits grassy stunt virus and had long been identified as a species with great potential for becoming a very serious pest. The brown planthopper is considered to be Indonesia's major insect pest and the greatest challenge for plant protectionists. When it was clear toward the end of 1986 that Indonesia was experiencing another set of brown leafhopper outbreaks, the government acted with great determination. First, it recognized that this pest was now resistant to certain pesticides and so the use of 57 biocide brands was outlawed and, second, the President announced that approved biocides were to be used only to control serious outbreaks, not as a preventative tool against possible pest attacks. In this way it was hoped that predator populations would be encouraged. There is, therefore, much greater government support at last for integrated ecological approaches paying attention to cultural controls (synchronized planting, etc.), natural enemies, pest surveillance, migration monitoring, different modes and patterns of insecticide use, and resistant varieties of rice. In short, the government will go to all lengths to ensure that the hard-won self-sufficiency in rice achieved in 1985 is maintained. The present situation shows just how relevant ecological approaches are to the management of agroecosystems.

Figure 9.5. Insect pests on different parts of rice plants at the early vegetative stage, late vegetative stage and mature stage.
After van Halteren 1979
The stem borers are caterpillars of pyralid moths which lay eggs on the young rice leaves. On hatching the caterpillar moves into a leaf sheath from where it bores into the stem. Depending on which part of the stem is bored into, the rice will either die or recover, but in the latter case will always produce a lower yield than an undamaged plant. Tryporyza innotata feeds only on rice. The rice leaf folder is also a pyralid moth caterpillar but this folds one or more leaves around itself and then feeds on the inside. This results in whitish streaks on the leaves and a badly infested field can take on a whitish appearance. Both adults and larvae of the rice seed bug damage rice by puncturing the seed with sucking mouthparts. This causes both physical damage and the introduction of bacteria and viruses which lead to both a reduction in yield and a reduction in grain quality. The areas heavily infested7 by the major pests in South Sulawesi vary between years in response to climatic conditions and effectiveness of biocides and other pest management (fig. 9.6). Some pests thrive on plants given high applications of nitrogen, others are favoured by the particular microclimate afforded by short varieties, while still others may be encouraged by a second crop of rice in a year (van Halteren 1979).

Figure 9.6. Changes in areas (000s ha) of rice fields in South Sulawesi during April-September, heavily infested with different pests.
Rats and their Control
As can be seen from figure 9.6, rats cause at least as much damage to rice crops as do the major insect pests. Rats cause the loss of at least 12 million tons of rice each year worldwide, and it is possible that rats and birds combined may have more serious impacts on rice crops than all other types of pests (Anon. 1976). Rats in Indonesia are probably responsible for a 5%-6% overall loss of the annual rice yield, but losses in individual fields can be much more than this. The rats live in tunnels they construct in the bunds between rice fields. These tunnels comprise main and secondary entrances and exits, blind tunnels for emergencies, and nest chambers.
The most serious rat pest in Sulawesi is the rice field rat Rattus argentivmter which damages rice crops throughout Southeast Asia, Two further species, the little rat Rattus exulans and the house rat Rattus rattus are generally found in peripheral areas but will enter rice fields to feed when other food is scarce. These species are usually only found close to buildings or farms.
Analyses of the stomach contents of rice field rats have shown that they all eat insects and snails, and that the amount of plant material they ingest depends on the type of habitat, and varies between species (Lim 1974). Rats eat rice at all stages of its development and although the theft of ripening rice grains is a direct loss from the harvest, the shredding of growing stems to eat the tender growing shoot does more absolute damage. A single, ripe seed head may represent a rat's daily food requirement, but the same rat may eat 100 growing shoots in a day before it is satisfied (Anon. 1976).
The reproduction of rats clearly follows an r-strategy (p. 345). The gestation period of the female rice field rat is 21 days, a litter generally numbers seven, and about eight litters are produced per female per year. Females could theoretically have more litters but reproduction tends to occur only when an abundant food supply is available, that is, towards the end of the rice-growing cycle. Male rats are capable of breeding at two months of age and female rats at just 1.5 months (Lam 1983). The mean life span of the rats is only about 4-7 months (Harrison 1956), but even by seven months a single female could have raised about 20 young.
The very high intrinsic rate of growth among rats is the major reason why attempts to exterminate them using traps or poisons are effective only locally or for short periods. If any rats are left, their litter size is likely to be larger than normal (up to 11) because competition for food is less and so the population will increase quickly. If, as rarely happens, all the rats in an area are killed, the unoccupied rice fields represent a wonderful opportunity for rats from other areas to colonize. If one thinks of an animal's niche as its 'profession', then when rats have been exterminated from a field, hundreds of 'jobs' become vacant for which the remaining rats are eminently qualified. It should be remembered that rats do not produce large numbers of offspring for the good of the species, or because that many are needed to fill all vacant 'jobs'. They are produced because rats are locked into their r-strategy and each female has to produce as many fast-growing, healthy young as she can, so that her hereditary line is not swamped by others. Many of the rats die young (as is shown by the life span figures above) but this is a consequence of the r-strategy. If the mother rat produced just a few offspring to whom she devoted her time for the fetching of food and for their defence, the available 'jobs' would have been filled by the larger number of other rats from other mothers.
Thus, control of rats in the long-term has to be centred on land management. Rats can live for only a few days on a diet of just rice stems (Anon. 1976) and this may be the only food at certain times, particularly if insect populations are also controlled in some way. Thus, if rice is planted and therefore harvested at the same time over large areas, and if scrub and other neighbouring habitats where rats could find alternative foods when rice grain was not available, were removed or utilized, rat populations could be kept within bounds. These are thus two of the main methods of rat control encouraged by the Department of Agriculture, the others being the digging out of rat holes and poisoning (Soekarna et al. 1980).
Attempts have been made recently at the national level and with the help of the armed forces to control rats but, despite the impressive piles of rats photographed for the newspapers, these will lead to only temporary improvements. Perhaps a more positive approach, as demonstrated in part of Central Java, is to consider rats as a resource which can be harvested on a sustainable basis and converted into animal feed.
Rice Field Birds
Among the more serious bird pests on rice are the Java sparrow Padda oryzivora, chestnut munia Lonchura molucca and the spotted munia Lonchura punctulata. The first of these was probably introduced into Sulawesi during the last couple of decades.
Birds generally range far wider and into more habitats than do insect pests and so their control poses considerable problems. Poisoning does not seem to be a viable control method because effective poisons are usually toxic against other animals, including man. The most common means of controlling birds is by scaring them with moving strings, cloth or scarecrows (fig. 9.7). In an account of bird pests, it is repeatedly stated that more ecological information is required for effective control programs (Adisoe-marto 1980).
Rice fields are also favoured habitats of various birds whose feeding habits suggest they may assist in keeping the populations of certain insect pests under some degree of control. The birds can be divided into two ecological groups: the rails and crakes, and the herons and egrets.
Rails and crakes are very secretive birds and are hard to see except at dawn or dusk when they are more active and come to the edge of thicker vegetation. They fly weakly, with rapid beating of short rounded wings and their legs hang limply below their bodies. They usually flee from danger by running or swimming back to the cover of dense vegetation. When walking they frequently cock their tails and they typically jerk their heads while swimming. Because of their secretive and possibly nocturnal habits little is known of their ecology but their diet certainly includes small insects.
The most conspicuous large rice field birds are the herons and egrets. The three major species are:
• The black-crowned night heron Nycticorax nycticorax (fig. 9.8) which, because of its nocturnal habits, is more often heard8 than seen.
• The cattle egret Bubulcus ibis, a white bird with yellow bill and greenish-yellow or blackish legs stands about 40 cm high. During the breeding season it develops a rufous-coloured head and neck. In Tana Toraja it is believed by many to bring bad luck if white birds are harmed and large numbers of these birds can be seen in the rice fields.9 There is a roost of over 300 of these birds in a bamboo grove by the road from Makale to Rantepao.
• The Javan pond heron Ardeola speciosa, a rather shorter and stockier brown bird with white wings. In the breeding season it develops a black back and a reddish brown breast but at other times it has brown and black speckled plumage.

Figure 9.7. An ingenious bird scarer from the beginning of this century.
After Sarasin and Sarasin 1905
The above species are rare or completely absent over many rice field areas of Sulawesi as a result of shooting, poisoning with biocides, or destruction of roosts. Both egrets and herons roost and nest in colonies and it is possible that these assemblies serve as information centres from which the members will gain knowledge of the location of the inherently patchy food sources (Fasola 1982). This is shown by the manner in which birds follow one another when flying from the roost to the feeding ground and the aggregations of birds foraging on the ground.
Although both cattle egrets and pond herons feed in rice fields, they do not compete with each other for food because they feed at different times and on different prey (table 9.3). Both birds are primarily insect eaters although the pond heron also takes frogs and fish, and the egret also eats spiders. The quantities involved are quite surprising. One Javan pond heron examined had over 500 individual prey items in its stomach, while one cattle egret was observed to catch 89 insects in just 15 minutes (Vermeulen 1985). However, neither these birds nor frogs appear to eat plant-hoppers or stem borers (Berry and Bullock 1962; Berry 1965; Yap 1976).

Figure 9.8. Black-crowned night heron Nycticorax nycticorax.

After J.W.C. Vermeulen pers. comm.
Grasshoppers are minor pests of rice fields in South Sulawesi and they are not regarded as troublesome on rice or any other crop in the Dumoga valley (K. Monk pers. comm.). One common genus Oxya has expanded pads on its feet which presumably help it to run across the water surface. The 3,000 cattle egrets resident in the rice fields between the villages of Toraut and Dumoga prey on an estimated 250,000 grasshoppers each day which must be a major factor in controlling their populations. Similarly, each day the 2,500 Javan pond herons in the some area probably prey on about 50,000 mole-crickets Gryllotalpa arachnoidea which are believed to damage rice roots. These are relatives of grasshoppers with reduced wings and an incredibly loud song made louder by the resonance provided by the tunnel from which they call.
COCONUT AND CLOVE
Coconut and clove are the two most important industrial crops10 in Indonesia after oil palm and rubber. Unlike the oil palm and rubber, they are both grown extensively throughout Sulawesi. North Sulawesi produces more cloves than any other province in Indonesia, and has nearly 10% of its land area under coconut. This compares with about 2% in Central and South Sulawesi, and 1% in Southeast Sulawesi (Bennett and Godoy 1986).
Dried cloves sell for about Rp 5,000-7,500/kg, and the yield from one hectare of mature clove trees can provide a smallholder farmer with Rp 5-10,000,000/yr.11 In contrast, one hectare of mature coconut palms provides a gross income of barely Rp 200,000. Mixed plantings of cloves and coconuts reduces the yields of both because of root competition and shading, but a farmer who intercrops his coconuts with cloves could increase his annual income to Rp 2,100,000 (Davis et al. 1985). During the 5 or 6 years before the clove trees begin to flower, he still has income from the coconuts which he can harvest every 3-4 months. The regular coconut harvests also provide income between clove harvests which usually occur annually, or sometimes once every two years for either physiological or climatic reasons which are not fully understood. This mixed cropping buffers the farmer against fluctuations in commodity prices and the effects of pest and disease outbreaks.
Coconut
The coconut is justifiably known around the world as the 'tree of life', 'mankind's greatest provider in the tropics', or 'one of Nature's greatest gifts to man'. The centre of origin of the coconut is probably in eastern Indonesia or Melanesia. The dispersal of coconuts to other tropical regions may have occurred naturally because the buoyant fruits can be carried great distances by ocean currents. However, the almost ubiquitous presence of coconuts on tropical sandy beaches owes a great deal to the movements of people (Purseglove 1968; Ohler 1984). Various parts of the coconut are used for food, drink, oil, medicine, fibre, timber, thatch, mats, fuel and to make utensils.
Coconuts can grow on a wide range of soils—sandy, coralline volcanic, alluvial, latosolic, lateritic and swamp soils (Ohler 1984). They require good soil aeration and drainage, and a constant supply of groundwater, which are all available in their characteristic location at the top of sandy beaches. Similar conditions are found in the volcanic soils. This is a major reason for the extensive planting of coconuts throughout Minahasa. Long dry periods adversely affect palm growth and nut production. For example, between July 1982 and April 1983 rainfall in Manado was just 369 mm compared to a previous average of 3,400 mm. Many coconuts died. It took about 2 years for nut production in the surviving palms to return to normal. This delay in recovery corresponds to the time it takes for a flower to develop and emerge from the heart of one palm and to develop into a mature bunch of nuts ready for harvest.
Coconuts were the world's major source of vegetable oil until the 1960 when soybean and palm oil became more important (Purseglove 1968). Coconut oil is principally made from dried kernel or copra which contains 65% oil. The oil was used primarily as cooking oil but the production from enormous coconut plantations in Minahasa, Toli-Toli and elsewhere provided the impetus for its use in soap and margarine manufacture in Europe during the later half of the 19th century. Secondary uses are in detergents, cosmetics, resins, wax candles, and confectionery.
Indonesia once obtained considerable export earnings from copra, reaching a maximum of US$31.4 million in 1970. Failure to replant ageing plantations established before Independence, together with an increase in the domestic demand for cooking oil since 1976, have led to a decline in the coconut export industry. About 60% of the cooking oil now uses oil from the oil palm Elaeis guineensis. It has been estimated at decreasing production in Central Sulawesi alone has cost Rp 6 billion over the last four years (C. Bennett pers. comm.). Since 1979, however, high-yielding hybrid coconut palms have been planted by farmers under the Smallholder Coconut Development Project. This hybrid, PB-121, is a cross between female Malaysian Yellow Dwarf and the male West African Tall.
A stand of mature coconut palms allows considerable light to reach the ground and multistorey intercropping can be practiced with coffee, cocoa, clove, bananas, sapota, pineapple, ginger, beans, maize or rice. Intercropping can both safeguard and consistently increase the income of coconut farmers considerably. The example of clove intercropping has been given previously (p. 588); another example is ginger which can produce 6,000 kg of rhizome/ha. At Rp 1,500/kg of rhizome, the farmer's additional gross income is Rp 9,000,000/ha. Coconut production is not reduced, but rather may subsequently increase because the palm benefits from the tilling, weeding and application of fertilizer for the ginger (Davis et al. 1985).

Figure 9.9. Characteristic 'V'-shaped notches caused by rhinoceros beetles eating the leaflets of the spear leaf.
A review of insect pests on coconut palms forty years ago listed 751 species of which 165 (22%) were not known to attack other plants. Among the best-known insect pests are the large, handsome rhinoceros beetles. The adults burrow into the spear leaves which consist of the unopened, folded young leaves. When these leaves open they have characteristic 'V'-shaped notches where Oryctes has fed on them (fig. 9.9). The beetle breeds in decaying organic matter, such as the pith of old coconut stumps, dung and decomposing leaves and husks. Rhinoceros beetle is a vernacular name given to two similar dynastic beetles: the adult male of Oryctes rhinoceros has a single backwardpointing 'horn' on the top of its head, whereas Xylotrupes gideon has two, forked, forward-pointing 'horns' (fig. 9.10). The first of these species is the more serious pest although even its effects are rarely fatal to the tree unless it reaches the single growing point beneath the terminal bud.
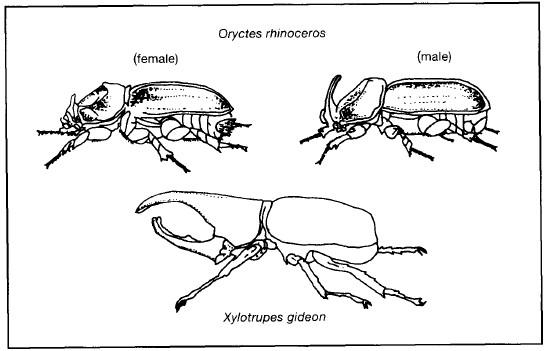
Figure 9.10. Rhinoceros beetles.
The palm weevil, Rhynchophorus ferrugineus, lays its eggs in soft or damaged parts of the palm crown. The larvae often burrow into leaf bases and the trunk through old Oryctes wounds. Attacks by the weevil are often fatal because their larvae usually destroy the growing point. Weevils and Oryctes can be controlled by the removal or burning of old trunks and their stumps. Biological control of O. rhinoceros using bacilovirus and Metarrhizium anisopliae has met with some success (Davis et al. 1985).
Coconut plantations in the Sangihe-Talaud Islands (and various locations in the Moluccas and Irian) suffer serious depredations from attacks by the long-horned (tettigonid) grasshoppers Sexava spp. with extremely long antennae (fig. 9.11). These 8-10 cm, green and brown insects seem to have very weak flight. This, in part, accounts for their absence on the North Sulawesi mainland. They lay most of their eggs in the soil at the base of the coconut palm. The eggs hatch in the evening and the first instar12 larvae climb the trunk and start eating the older leaf blades. The larvae generally spend the day in the canopy and the night on the ground. Infestations are more severe on older leaves, possibly because of the toxic trichomes on the very young and expanding leaves. Trichomes are minute hairy outgrowths concentrated on the lower surface and edges of the leaves. They contain chemical defences and may also discourage insect attack simply by physically obstructing their access to the leaf blade itself. Defoliations of coconuts on Sangihe-Talaud can be so severe that whole communities of smallholder farmers have had to abandon their coconut gardens and move to other areas (Kalshoven 1981; Davis et al. 1985).
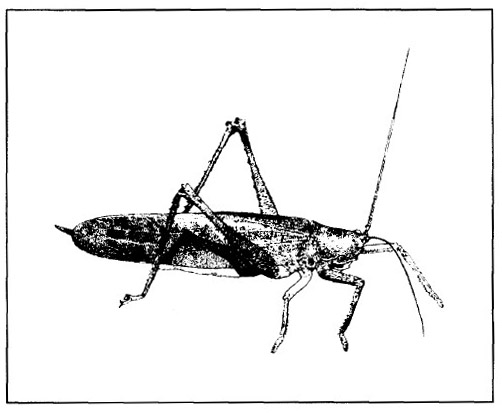
Figure 9.11. Sexava long-horned grasshopper.
Trichomes apparently also discourage leaf-mining hispid beetles Plesispa reichi and Brontispa longissima (fig. 9.12) (Davis et al. 1985), but serious outbreaks of the latter have been recorded in all provinces of Sulawesi. They attack the spear leaf particularly on young palms and are most abundant during dry periods. Plantations in South Sulawesi in the 1920s and '30s were brought under control using the eulophid wasp Tetrastichus brontispae as a parasite. The wasp can attack 60%-90% of the pupae and 10% of the larvae; about 20 wasps will emerge from a single beetle pupa. This parasite was also successfully released in North Sulawesi including the Sangihe Islands. Plesispa reichi is a blacker relative distinguished by a much darker carapace. It is generally found on seedlings and young coconut palms.

Figure 9.12. Hispid beetles.
Nettle and slug caterpillars (Cochlidiidae/Limacodidae) are amongst the most serious pests, causing considerable damage to coconut palms and other crops in Southeast Asia. They are easily recognised by their gelatinous bodies and slug-like appearance. Some species have colourful, barbed spines on their back and sides. If touched, these barbs enter the skin and release a toxin which can elicit an acutely painful allergic reaction. The brilliant colouring of the nettle caterpillars, as in many insects, warns potential predators of the unpleasant consequences of an attack. In contrast, the adult moths are often small and dull coloured. In 1978 one widespread species Setora nitens devastated large areas of coconut plantations throughout Indonesia. Observations at the Coconut Research Institute in Manado showed that the dwarf palms suffered less than tall varieties probably because of the higher density of leaf trichomes in dwarf palms. Some farmers reacted to the outbreak by pruning the infested leaves but this led to young fruits being scorched by the sun and to flower shedding. The pruning led to a 76% loss in yield compared with a 14% drop for unpruned palms. In the same year there were serious outbreaks of Chalcoscelis, Darna and Parasa nettle caterpillars. Individual coconut leaves were attacked by as many as 700 caterpillars at once.

Figure 9.13. Parasa balitkae in characteristic resting position.
Darna catenata is a Iimacodid species apparently restricted to Sulawesi (including Muna and Sangihe) and Irian Jaya. It is found primarily in dry areas and is an important pest around Palu (pp. 484 and 487), where outbreaks seem to be triggered by drought. One major outbreak occurred there in 1950 when more than 100,000 coconut palms were badly damaged (Metzner 1981). In 1982 the same area was devested and 700 ha of palms appeared to have been scorched by fire. By 1984 an additional area of 9,000 ha was affected.
A new coconut pest species which is locally important, Parasa balitkae13 has recently been described from North Sulawesi (fig. 9.13). This and the other nettle caterpillars are parasitized, sometimes with 100 wasps of several species and families emerging from a single caterpillar. These have considerable potential as control organisms.
One of the most destructive coconut pests is the endemic pig Sus celebensis, which uproots newly-planted seedlings in order to eat the tender heart or cabbage. Other mammals, including rats, may cause as much loss to the coconut grower as insect pests, but fewer resources are devoted to their control. Rats live in the crowns of coconuts and are a persistent problem because they damage young leaves and nuts of all ages. They can also consume stored copra. Rats can cause losses of 20% and occasionally as much as 50%. Rat poisons have been used with some success but rat resistance and cunning have reduced their efficacy.
It is very unlikely that rats could ever be exterminated and so a major aim should be to keep populations as low as possible using sustainable and natural means of control. One method used in some oil palm plantations is to encourage the widely-distributed barn owl Tyto alba to establish its territory among the palms. Analysis of pellets of bones regurgitated by barn owls showed that these birds feed almost exclusively on wood rats Rattus tiomanicus, and an adult pair of owls could be expected to kill 1,300 rats/yr within a territory of about 20 ha (Lenton 1983). In oil palm plantations the wood rat commonly lives at a density of 250/ha. Thus an owl territory could contain a standing crop of 5,000 rats, one quarter of which would be killed each year. It should be remembered, though, that the total number of rats living in an area during a year would be greater than the standing crop because the rats are continually reproducing and dying. A study of owl predation in England found that 20%-30% of the population of mice was taken every two months. The barn owl T. alba is not found in Sulawesi, but the grass owl T. capensis occurs in Central and Southeast Sulawesi, the endemic Minahasa barn owl lives in North Sulawesi, and the endemic Rosenberg's barn owl T. rosenbergi in all provinces. These owls have not been reported from coconut plantations but they could possibly be encouraged if nest boxes were erected at 6 m on electricity, telephone or other poles (fig. 9.14), because nest sites appear to be a limited resource (Lenton 1984). There is no guarantee that these owls, which typically inhabit forest rather than open areas, would adopt these nest boxes, but they are cheap to make and erect and their low cost would compare very favourably with control programmes using chemical poisons. A simple pilot study would show if this were a useful course to pursue.
The territory of barn owls in oil palm plantations appears to be a minimum of about 20 ha. Territory sizes are determined, in part, by abundance of essential resources but there are also strict upper and lower limits. The minimum size of territory is probably determined by the area required during periods of lowest prey density and the maximum by the area that can be effectively monitored and defended without diverting too much time and energy from other important activities. The population of rats in a coconut plantation is likely to vary through the year, but the resident population of owls would not continually shift territorial boundaries. Once established territories are probably quite stable, and the owls respond to fluctuations in prey density by producing more or less young. Their breeding strategy appears to be to produce large clutches and broods as frequently as possible in habitats with high potential carrying capacity (Lenton 1984).
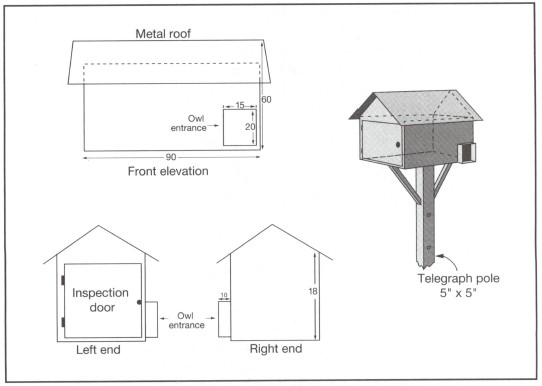
Figure 9.14. A nest box likely to attract owls to coconut plantations to control rats. The box dimensions are 0.5m x 0.5m x 1 m. Numbers in above illustrations indicate measurements in centimetres.
After G. Lenton pers. comm.
Diseases are not generally a problem in coconut palms, but some coconut diseases are locally important. One of these is premature nutfall disease outbreaks of which are caused by the fungus Phytophthora palmivora and are correlated with the peak of the rainy season in December to January (Bennett et al. 1985). Another disease affecting the leaves is often caused by another fungus Pestalotiopsis palmorum invading leaf wounds made by the limacodid pests.
Clove
Clove trees Syzygium aromaticum (Myrt.) are indigenous to the Moluccas and for more than 2,000 years the dried, unopened flowers (the cloves) have been used in east Asia as a spice, as a cure for tooth decay and as a breath sweetener. Indonesia is the world's largest producer, consumer and importer of cloves. Numerous parts of Sulawesi have caught what is often referred to as 'clove fever'. In areas such as Toli-Toli, hillside after hillside is covered with a regular forest of clove trees. Those people who own cloves receive a good or excellent income.
Indonesia consumes 25-30,000 tons of cloves each year of which 10%-30% is imported, primarily from Zanzibar. About 85% of dried cloves are shredded and added to tobacco in the ratio of about 1:2 to make 'kretek' cigarettes. Kreteks were already being smoked in the Moluccas at the start of the 17th century (Alauddin 1977). North Sulawesi produces 29% of the Nation's home-grown cloves, followed by Central Java (18%), Lampung (15%) and West Sumatra (13%).
The clove tree grows best at low altitudes where there is a marked dry period. The annual rainfall in its natural habitat is 2,000-2,500 mm and the agroclimatic zone is 'D' or 'E' (p. 22). Good drainage on deep acid loams seem to be important, as is shade in the early stages of growth (Purseglove 1968). Clove seeds remain viable for only a short length of time, and all these characters reflect its origins as a small (<14 m) forest tree unable to germinate and grow in full sun and unable to wait for favourable conditions (p. 359).
Pests of clove trees include a number of longhorn beetles (Cerambyc-idae). For example, the larva of Nothopeus fasciatipennis burrow into the trunk through a 3 mm x 5 mm hole which eventually seals itself with a white calcareous crust. The larvae tunnel both in the heartwood and just below the bark. Where these tunnels extend right around the trunk they may ring girdle the tree and kill it. A tunnel may be several metres long. The wasp-like adult (fig. 9.15) emerges higher up the tree through kidney-shaped holes.
Other groups of borers include moths of the family Metarbelidae whose larvae live in tunnels in stems, often near injuries, and feed on bark. It was estimated recently that 30,000 of the 5 million clove trees in Toli-Toli are infected with these stem borers. They pose a considerable threat to clove production in Central Sulawesi since there are only another 2 million clove trees in the province (Anon. 1986b). Scale insects (Coccidae) are quite common on cloves but they do not have serious deleterious effects. Many of these pests are controlled successfully using biocides. Mammalian pests on clove trees are restricted to squirrels which chew bark off the young twigs.
Most serious of all is a disease which was first detected among cloves in the Sonder region of Minahasa, and which is known simply as 'clove leaf drop'. Although it has been suggested that it is result of inappropriate methods, its pattern of spread is not consistent with this. The disease appears to be triggered or exacerbated by drought but the causal organism and its mode of action are still unknown. Leaf blight associated with a fungus Phyllosticta sp., has recently affected hundreds of hectares of cloves in Tana Toraja to which it was introduced from either Sumatra or Java. Still unknown is the cause of Sumatra disease which was first recognized with certainty in 1961 in West Sumatra where it may have been present since the 1920s. The disease is caused by a xylem-limited bacterium (Bennett et al. 1985). It is now found in North Sumatra, West Sumatra, Bengkulu, Lampung and West Java. Thousands of hectares of clove trees have been destroyed by the disease and its appearance in Sulawesi would spell ruin for many people.

Figure 9.15. Nothopeus fasciatipennis.
COTTON
Cotton Gossypium hirsutum (Malv.) is the world's major fibre crop and covers about 30,000,000 ha or 5% of the world's cultivated land area. Conversion of land to cotton can bring considerable economic benefits.14 particularly in Indonesia where textile exports are so important, but there are a number of potentially serious negative impacts on the environment, the causes of which need to be continuously monitored.
Cotton has been grown in Indonesia for many years but interest started to increase during the Japanese Occupation at the end of World War II. In Sulawesi, cotton is grown by the State Plantation Estate Company XXIII in South Sulawesi, and by P.T. Kapas Indah in Southeast Sulawesi. The second company runs a nucleus estate and small holder project near Tinanggea which began in 1977. It currently covers an area of about 5,200 ha, of which the mechanized nucleus covers 1,200 ha, and 4,000 ha is operated by smallholder, contract growers. The nucleus used to cover 1,700 ha but 500 ha were turned over to producing hybrid coconuts because the competition from American cotton is so great (Moeharram pers. comm.).
The first obvious danger is that, for any crop grown intensively over large areas, pests can wreak havoc and financial disaster. To counter this it is tempting to use large quantities of biocide and it is revealing that more biocides are used worldwide on cotton than on any other crop (Goodland et al. 1984). The problems encountered by pest control programs are well illustrated by the experience of P.T. Kapas Indah. It seemed unlikely that a new crop in a new area would suffer much from pests, but the entire crop of the first year was wiped out. A leafhopper Empoasca found the crop early on and reached such alarming densities that a major and successful spraying programme was undertaken. Unfortunately, however, it seems as though this programme exterminated insect predators and what remained of the cotton crop was destroyed by the earworm Heliothis armigera, which is the caterpillar of a noctuid moth which could not be controlled with the chemicals available. In the second and third years the number of biocide sprayings rose from between 3-5 to 5-7 respectively. In the fourth year, even though the crop was sprayed thirteen times, almost all the crop was lost to Heliothis, largely because of unexpected rains which prevented the biocide from reaching its targets. Over the following three years, the biocides used were changed and the sprayings were reduced from seven, to six to five. For the crop just harvested (1985-86), however, seven sprayings were necessary, presumably due to pest resistance, and next year the biocides will be changed again (R. Iskandar pers. comm.). Interestingly, the smallholder fields consistently suffer less insect damage than the fields of the nucleus estate and this is probably due to the position of the cotton between other crops and the dilution of the cotton in its field due to intercropping with other plants (M. Wade pers. comm.).
The control of cotton pests needs to be based on integrated control programmes that take into account the ecology of the pests and their enemies. For example, it is common practice to have four to six months (during the dry period) with no cotton crops in the fields in order to break the breeding cycles of pests. It is less common for surveys to be conducted to determine what other plants the pests may be using as alternative foods for the larvae. The growing plants need to be monitored for eggs and larvae of pests but their presence does not automatically require the initiation of a spraying programme. Many crops can tolerate heavy (<50%) loss of their leaves while young, and early spraying may retard the development of pest predator populations.
The development of resistant crop varieties would undoubtedly be the best approach (hairy varieties of cotton are, for example, resistant to Empoasca), but until these are developed for the pests of cotton in Sulawesi, the best approach is to maintain a range of biocides with different strengths to counter a range of pests under different conditions. The use of the biocides would be determined by regular monitoring of both pests and predator populations. For example, at P.T. Kapas Indah, the threshold of pest density is an average of 1.5 larvae per 10 cotton plants. Only when this density has been found will spraying be allowed.
Cotton has the potential to be quite lucrative for the producers. It will be viable, however, only if sound pest management can be introduced that reliably minimizes pest damage while safeguarding the profitability of the project and the quality of the environment.