7
Mimicry is a form of expression that one might not imagine to feature in an exploration of cognitive complexity in birds. After all, when parrots ‘parrot’, they were thought to mindlessly follow patterns that they have heard, but it is not that simple. Mimicry may be mindless or the first step in learning. There are two behavioural forms of mimicry that are of particular interest here because they may have a cognitive dimension: one is mimicking another species’ vocal performance and the second is mimicry of motor movement. Mimicry of movement is rare (or has not been described in detail) and was thought to be unique to humans, but it was the budgerigar and the blue tit that showed this belief to be incorrect. This will be discussed later in this chapter.
Mimicry can take quite a number of forms. Some have to do with mimicry of shape or colour of a species (i.e. a harmless species mimicking another poisonous species by having evolved a similar colour or, sometimes, shape). Entomologists developed very extensive models of mimicry long ago and, at least in insects, such a relationship is premised on honest and dishonest signals to either secure prey or to avoid being eaten. This is known as Batesian mimicry. True mimicry involves three parties: the true identity of the mimicked one, say butterfly A; the mimicker, called butterfly B; and the predator that is deceived by butterfly B, i.e. the predator will not eat B because it looks like the unpalatable butterfly A (Wickler 1968). However, this traditional model is not really translatable to vocal mimicry in birds, although attempts have been made to apply the model of Batesian mimicry to an avian species (Veerman 1994). Batesian mimicry also has nothing to do with cognition.
Vocal mimicry
Vocal mimicry in birds generally involves only two parties and the mimicking bird has not evolved to always appear (vocally at least) as if it were another species (as have look-alike insects). But the topic is more complex than this. Vocal mimicry, according to statements of some 40 years ago, was defined as the imitation by birds of sounds outside their species-characteristic vocalisations (Thorpe 1964). Armstrong (1963) similarly argued that mimicry is ‘the copy of sounds other than those of the bird’s own species’. However, this definition may not encompass all forms of mimicry. First and foremost, birds may mimic their own parents or members of their group. Indeed, such copying, as has recently been confirmed, is important for communication. Being absolutely accurate about a specific sound profile actually facilitates and enhances communication within groups (Wheatcroft and Price 2013). In addition, birds may copy sounds of neighbours of the same species (Putland et al. 2006), closely related species or may take snippets or segments from other avian species into subsong, but not full song, or they may copy even mammals or the sounds of inanimate objects, such as car horns or chainsaws, rather than that of other species (Gramza 1972).
The puzzling part about mimicry is that, on the one hand, bird vocalisations are species-specific – and, indeed, they are among the most robust ways of identifying a species – while, on the other, the performance of mimicry undermines the very concept of species-specificity (Hausberger et al. 1991). By definition, mimicry is not part of the species’ repertoire (or genetic template) and there may not even be any natural propensity for the mimicked sounds in that bird’s own repertoire (Kelley and Healy 2011). For instance, mimicry of dog barking is far removed from a galah’s or a magpie’s natural repertoire and yet I have records of both giving believable renditions of dog barks.
In other words, mimicry seems an unusual vocal expression. Its existence is nevertheless of great interest now, especially to neuroscience and cognitive science. Better than almost any other form of vocalisation, it can demonstrate very well that a bird uttering clearly identifiable mimicked sounds must be capable of learning. Indeed, mimicry is the clearest possible evidence of vocal learning. There is no doubt at all now that examples of mimicry may be an excellent way of exemplifying and analysing vocal development and learning (Greenlaw et al. 1998). In fact, vocal learning often consists entirely of imitation. Thus, the line between song learning and ‘artefact’ (be it in captivity or in the wild) has become rather blurred.
Parrots can imitate the vocalisations of other birds, are excellent mimics of human speech and general noises, such as doors closing or telephone ring-tones (Baylis 1982), and their repertoire in captivity can be substantial. However, so far, only very few examples of vocal mimicry by parrots in the wild have been reported anywhere in the world. One is that of the African grey parrot (Cruickshank et al. 1993). In other isolated cases, in Australia, there is a reported case of pink cockatoos raised by galahs adopting the vocalisations of their host parents (Rowley 1990) and a yellow-tailed black-cockatoo was once heard apparently mimicking a kookaburra (Sharland 1938).
Apart from parrots, there are many species mimicking in the wild. Alec Chisholm was the first to list 56 Australian avian species as having some ability in vocal mimicry in the wild (Chisholm 1948). Since then, about half of the ones he named have been verified. Mimicry is thus surprisingly widespread among Australian birds and in some verified cases even highly developed. Few people know that spangled drongos (Thorogood 1941) are good mimics and so are bowerbirds (Frith and Mcguire 1996), including the satin bowerbird (Rowland 2008) and regent bowerbird (Chisholm 1951). Australian species that mimic include members of the Cracticidae, such as magpies (Kaplan 2000), currawongs and butcherbirds (McDonald 1940), some honeyeaters and many others, such as the speckled warbler (Courtney 1967), Richard’s pipits (Bourke 1943), brown thornbill (Hindwood 1933), southern figbird (Holland 1962), and Horsfield’s bushlark (Cameron 1936). The olive-backed oriole has also been reported to mimic in the wild (Beland 1977). Occasionally, there are reports of mimicry that turn out to be incorrect. For instance, there was a report on mimicry by a tawny frogmouth (Legge 1934), but both sounds described (one that was similar to the ‘oom’ of a powerful owl, the other of the raspy part of the kookaburra laugh) are conceivably rare vocal expressions within the tawny frogmouth’s own repertoire. My own research on tawny frogmouths over a 10-year period does not support the observation of mimicry (Kaplan 2007a).
The question some researchers have asked is whether the importance of vocal imitation has not been exaggerated. Vocal mimicry has always been considered marginal to science in that it seemed to have no particular function (Robinson 1975). Any interest in vocal mimicry was generated first by the early assumptions of the instinctual basis of species-specific behaviour, of which vocalisations were thought to be a part (Witchell 1896). According to West and King (1996), vocal imitation may, in some cases, be no more complex than a reflex action and thus may be among the most rudimentary of imitation behaviour. They agree with Greenewalt (1968) that, far from elucidating important links between the ability of vocal learning and cognitive capacity, production of acoustic ‘photographs’ of sound may be an impressive motor skill but may say little about cognitive and social consolidation of communicative behaviour (West and King 1996). Thus, mimicry, particularly by various psittacine species, such as budgerigars, cockatiels, galahs, corellas and sulphur-crested cockatoos, has been discredited as an artefact of captivity and hence as being of no further scientific interest.
But do birds merely ‘parrot’ and is it just an artefact of captivity? Irene Pepperberg tried to convince us otherwise by engaging in meaningful vocal exchanges with her African grey parrot, Alex (Pepperberg 2006c). And indeed, vocal imitation cannot be so easily dismissed, because it is a widespread behaviour, especially so in Australian birds. Vocal mimicry is a rather special ability performed by psittacine species, songbirds, cetaceans, by some seals and humans (Moore 1996). Indeed, it may well be classified as an extremely complex type of behaviour, as Gramza (1972) noted, occurring only in very specific classes of vertebrates.
Among European birds, the starling (Hindmarsh 1984; West and King 1990; West et al. 1983) is the star among mimics. In North America it is the northern mockingbird (Gammon and Altizer 2011) and the brown thrasher, a songbird with a considerable repertoire, that is also known to mimic (McAtee 1940). In Australia, the lyrebird and the Australian magpie vie for first place of the most remarkable songbird and best mimics in Australia, if not the world.
For cognitive sciences, vocal mimicry is exciting because its existence provides a useful tool for measuring such incidents and can deal with a universal question of how vocal learning is achieved. The acquisition of new mimicked snippets requires a receptive brain and memory. It requires a brain that is ‘plastic’ enough and has the capacity to acquire new sounds not part of its species-specific repertoire, has special auditory feedback mechanisms and the possibility of storing sound as memory, plus a vocal apparatus that can produce the mimicked sound (discussed below).
The context of mimicry
Mimicry may occur within the breeding song, and another within territorial song. The best examples of either are the songs of magpies and of the lyrebird.
In songbird species in which the male sings to attract a female, as in zebra finches, juvenile males copy the song of their fathers (Catchpole and Slater 2008), but then the song ‘crystallises’ and remains more or less unchanged for the rest of the bird’s life. Songbirds with these characteristics are thus generally not candidates for mimicry. Compared with the many high-latitude species with these characteristics, this category of bird tends to be rather underrepresented in Australia, while birds with the potential to mimic is substantial. This has several reasons. One, as mentioned before, is that competitive songsters are polygamous males who leave the parenting entirely to females and such species form a very small minority among Australian birds. Another reason is because some birds need to retain some plasticity throughout life, even if they have a territorial song, if only for the purpose of tuning in to neighbours (Putland et al. 2006) to indicate their belonging to a territory or region, and that may change several times during their own lifetime (Kroodsma et al. 1982). Finally, some birds that are improvisers, as magpies are, never have one specific and ‘fixed’ song, leaving plenty of room for mimicry because the brain remains plastic to process new vocal stimuli and can memorise and reproduce them even when not species-specific.
The best-known species for mimicry in the wild are lyrebirds of both species (superb and Albert’s), and Australian magpies of any subspecies, Australia-wide. They are the most accomplished vocal imitators and they both have a large mimicry repertoire. The lyrebird seems to contradict the previous paragraph. Lyrebirds are indeed polygamous and promiscuous and they fertilise several females in a season (Schodde and Mason 1999). They take no share in nest building, incubation or in rearing the young. A 7-year study found that, among the vocalisations of superb lyrebird groups studied (seven in all), there was structural, locational and temporal constancy of territorial song from one generation to the next (Powys 1995) suggesting that male youngsters learn the much simpler part of vocalisations (territorial song) from their fathers after fledging. And yet they mimic and, even more dramatic is the fact that their breeding song is not even their own! All the pain of tuition that polygamous males generally undergo just to learn their song perfectly, and here is the lyrebird that plays the game quite differently. The lyrebird retains a plastic brain and then proceeds to copy – not all at once, but from season to season – expanding its repertoire until it has a most impressive performance together that may impress the females. All this is something that is outside the expected and generally known parameters, as so many things in Australian birds seem to be. Indeed, the entire and dramatic male lyrebird dance on a mound in the forest – all those loud vocalisations that can be heard for miles – consist almost exclusively of mimicry of other bird species!
Lyrebirds appear to string mimicked sequences together in an unchanging order, referred to as the Albert cycle (Curtis 1972; Robinson and Curtis 1996). The male picks a few mimicking seconds and uses these snippets like a pearl on a string, adding to that string as he sees fit. Additions are made to that string and, when he has come to the end of that sequence, the bird eventually returns to the beginning of the sequence and starts all over again in the same order. Obviously, the longer the string, the more dazzling and varied is his performance. Favourite borrowings from other species for his own song are the ‘laughing’ (i.e. staccato, parts of the kookaburra calls), the screeching sounds of the yellow-tailed black-cockatoo, the contact calls of the currawong and, in the Dorrigo National Park to mid-north coast forested areas, lyrebirds have added the eastern whipbird, the golden whistler and the catbird to their favourite repertoire. Snippets of many other species may be added along with sounds belonging to inanimate objects that may add specific percussionist elements of surprise, such as the sound of a car horn or a chainsaw. A documentary hosted by David Attenborough and filmed in Melbourne’s Mt Dandenong area (Sherbrook Forest) even documented the sound of a camera release button performed by a displaying lyrebird male.
Mimicry in the lyrebird is thus just for show to impress a female and to dazzle her both with his ‘lyre’ (the two special tail feathers that get shaken while he dances) and his song. His song is unique in the world and a tour de force all its own.
In addition to song and dance, the Albert’s lyrebird adds a rhythmic pattern to the display by shaking nearby vines (Curtis 1972) as if to accentuate the performance at rhythmically emphasised moments. This is a theme that has been newly picked up and it is finally recognised as an unusual ability and recognises Curtis’ work 40 years after his discovery (Dalziell et al. 2013). Importantly, all the mimicked parts are learned parts. Not only will he have to memorise the sounds, he will have to remember the sequence of all the mimicked segments and will need to keep that memory from one year to the next because, in between breeding seasons, males and females produce only a brief territorial song.
The role of mimicry in magpies is far less clear because it cannot be tied to a function such as territorial defence or to the breeding season. In magpies, mimicry is a small subset of sounds within an extraordinarily large repertoire and it occurs only within the context of one type of song that is generally referred to as warbling, but it seems to be randomly distributed in song sequences (Kaplan 2000) and is performed both in solitude and in company, independent of the breeding season and is voiced by males and females alike (Kaplan 2008).
David Attenborough claimed in his series ‘The Life of Birds’ (BBC, 1998, Episode 7) that lyrebirds knew about 20 mimicked sounds. For magpies, 28 types of unambiguous mimicry were found (see Table 7.1) in 10 years of searching for and analysing magpie mimicry (Kaplan 2000, 2005). The magpie, unlike the lyrebird, also copies a range of mammal sounds: foremost dogs and horses (Pollard 1930; Conway 1940; Lord 1941; Thorogood 1941; Chisholm 1946, 1947; Boehm 1948; Collins 1983; Sanderson and Crouche 1993). Magpies also produce imitations of the human voice and can whistle in an almost human fashion (Chisholm 1937; Sanderson and Crouche 1993; Kaplan 2000). Imitations of the human voice means that the birds produce individual human words but also entire sentences and thus may raise the number of items of vocal mimicry considerably above the 28 that were identified as part of a rigorous study of magpie vocalisations (Kaplan 2005) (Table 7.1). My recordings include phrases such as ‘Go away’ and ‘I’ve got dinner for you’ and a magpie, very believably, mimicking horse neighing. Those renditions have been enjoyed by many audiences at international conferences, always with an air of disbelief at the precision of these renderings. Below (Figs 7.1–7.2) are just two examples provided as sonograms. (These multiple recordings of mimicry do not refer to repeat bouts of the same mimicry by the same bird but to evidence of the same type of mimicry by different birds.)
There is only one reported case in which transmission of mimicry was apparently passed on from one magpie to another. According to Waite (1903), a captive bird was taught a tune played on a flute, and a second bird learned the tune from the first bird, the two performing it as a duet: when the first commenced the tune the second came to attention, and with half-open bill waited for the first bird to finish its part, whereupon it continued with the remainder. There has been no other report of this kind (Kaplan 2005, 2009). After I had collected over 80 continuous hours of magpie mimicry sounds (probably the largest sample yet collected) the notion that mimicry may be no more than a mistake in copying their own species-specific song, as Catchpole and Slater (2008) had argued, had to be dismissed. The accuracy of the magpie’s mimicked sounds leads one to the conclusion that this species has a very highly developed auditory perception and great musical discriminatory abilities. However, all this is ultimately saying is that basic qualities can be developed and enhanced through learning. The fact that both magpies and lyrebirds need to practise to retain the mimicked items should lead one to assess the mimicry in a positive rather than negative light: not as a mistake but as an acquired capability.
Table 7.1. Species identified positively as being mimicked by magpies
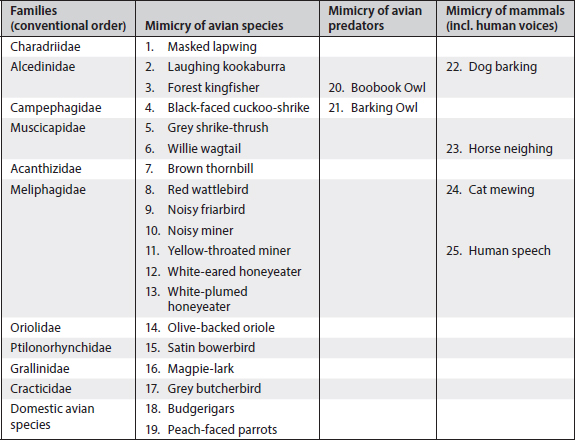
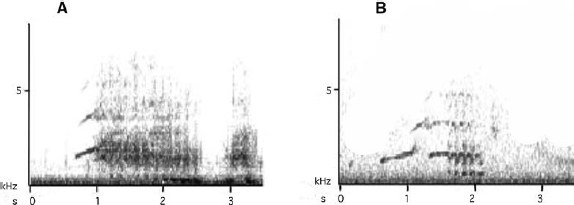
Fig. 7.1. (A) Sonogram of sound recording of horse neighing. (B) Magpie rendition of horse neighing. The y-axis is the frequency in kHz; the x-axis time in seconds. The black areas in panel A represent noise. Note that the magpie has faithfully copied the frequency (including overtones at 4 and 5 kHz) and the length of the bout.
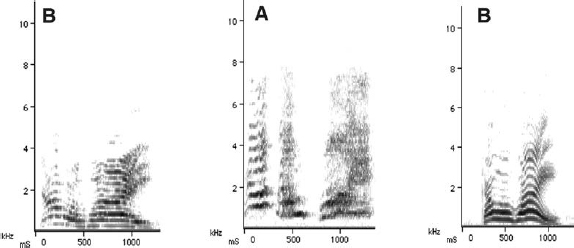
Fig. 7.2. Magpie mimicry of the human voice. (A) The mimicked rendition of the phrase ‘go away’. (B) Two different human speakers also spoke the words for comparison, but linked the words differently. (Left) In the first human speaker, the separate word ‘a’ is pronounced separately; (right) in the second speaker, the ‘o’ and ‘a’ are slurred together. The magpie had learned the phrase from either speaker but the bird had clearly decided to separate the words very precisely. This rendition is louder than the original but the temporal patterns are again correct and the harmonics mimic the human voice very well. (From Kaplan 2005.)
Functions of vocal mimicry
Quite a few papers have been published based on the theoretical assumption that mimicry may have functions that are adaptive (Baylis 1982). In some cases, function can even be identified although it is unclear in most cases of mimicking by Australian birds. One function is embellishment of song in order to impress females, as already described in lyrebirds (Robinson and Curtis 1996; Thoburn 1978), and also described in the regent honeyeater (Veerman 1994).
Another category of potential function may be the outlining of a vocal map of a territory. The only bird sounds mimicked by magpies were of species that permanently shared their territory, suggesting selectivity as if to recollect the acoustic characteristics of its territory. Mimicry by magpies may thus indeed be adaptive).
Mimicry and cognitive complexity
There have been comprehensive reviews of mimicry in recent years (Kelley et al. 2008; Wickler and Hauber 2013) indicating that this topic is not going away. It has also been known for some time that mimicry may well give us valuable clues about song development (Marler and Mundinger 1971; Marler 1991). Hauser et al. (2002) noted in particular that the ability to imitate (vocally and otherwise) was a crucial prerequisite of the ability to use language as a communicative system (see also Chapter 9). Suffice it to say here that the large vocal repertoire of magpies may be an indication of a basic ‘lexicon’, not homologous to human speech but an efficient system to enhance social living and survival as a group. Mimicry may serve an essential computational aspect of memory formation, but this in itself would be functionally adaptive. In lyrebirds, the function is clear, but the feat of memory is still remarkable, although not entirely unusual.
However, a crucial breakthrough in neuroscience shifted the topic of vocal, as well as movement, imitation (Moore 1992) from being of marginal interest to centre stage when the underlying processes in imitation were discovered. Indeed, advances in avian brain research between 2004 and 2008 changed the field in such fundamental ways that it is almost impossible to conceive of how we thought before they were made. In 2008, Prather and colleagues discovered ‘mirroring neurons’ used for learned vocal communication. They showed convincingly that a bird listening to a conspecific’s vocalisation activates neurons that would also fire had the bird sung the same passages itself (Fig. 7.3).
In other words, a specific set of neurons responded to primary songs equally when the bird sang them and when it heard them, fulfilling the criteria for mirror neurons identified in primates a decade earlier (Di Pellegrino et al. 1992; Rizzolatti et al. 2001; Umiltà et al. 2001) and in humans recently (Keysers and Gazzola 2010).
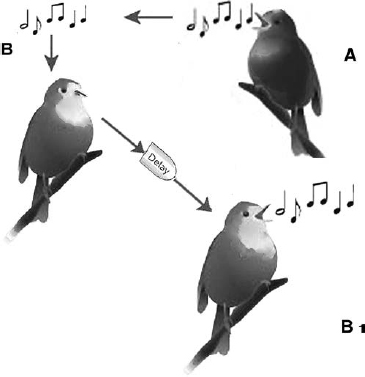
Fig. 7.3. Acquiring sounds by vocal imitation. Drawing based on Tchernichovski and Wallman (2008). A diagrammatic representation of a whistler in the process of acquiring sounds by vocal imitation. Bird A sings, bird B listens but does not sing. The passage will be reproduced unchanged by bird B (in B1) after exposure of only a few seconds it took bird A to sing the sequence. There is a known delay in reproducing any learned vocalisation of up to 48 h but thereafter it is copied in its entirety (Prather et al. 2008).
The way it works, as Tchernichovski and Wallman (2008) explain, is that, on input, the motor signal is delayed and this implies that the mirror neurons are providing a ‘corollary discharge’ signal: that is, a neural representation of the song being heard is available to the bird that first heard it and can now check that encoded version against the song it then sings. Or expressed differently, the bird gets a copy of the song in its memory against which it can judge its own output (performance) of the song. Importantly, the mirror neurons identified by Prather et al. (2008) belong to a population of neurons that is not replaced, as other neurons in the song system are, but is stable across song development. It is this stability that enables the juveniles to learn by copying.
In summary, one outcome of the discovery of mirror neurons has been that mimicry has moved out of the old discredited category of the ‘mindless’ and insignificant. Mimicry has been propelled to the status of a phenomenon important enough at least as the first and crucial step in learning. The mirror neuron discovery has thus turned listening into learning and imitation into a very important part of learning (Byrne 2009) and it explains how birds, even if not tutored, can learn song (Fig. 7.4).

Fig. 7.4. Juvenile learning by listening. Here is a rare example of an active listening act in a juvenile magpie (in front, adult female parent behind, carolling). The juvenile stands completely still during the carolling bout of the parent, raises its head to about the same angle and places its legs in a slightly angled position. Carolling bouts are of great amplitude and take substantial energy and can only be produced when the head is moved backwards (details of postures during singing are explained in great detail in Kaplan 2005). The juvenile has adopted the same posture as the parent bird. There is no sound as juveniles are not permitted to participate in territorial calls but the postural changes and particularly the plumage extension on abdomen and at the neck (typical when magpies sing) indicate that the musculature used for song is in use and that the bird is listening and imitating at the same time. It is passive participation in the form of intense listening.
Celia Heyes (2009) summarised the interest in imitation and mimicry very accurately and, although this was meant to consider issues in human development, it might as well have been said of birds. She said:
Imitation is an important and intriguing neurocognitive process: a process that bridges the gap between one mind and another; that powers cognitive and social development in infancy and childhood; that promotes empathy, cooperation and well-being in our relationships with others (Heyes 2009).
Although there are many theoretical and difficult debates when trying to marry neuroscientific research with learning outcomes, and when jumping from cellular level to social level (Hickok 2009), it is undeniable that mimicry and imitation are now respectable categories for investigation. They are international and multidisciplinary and the entire question of the progression of learning is now often tested in the context of mimicry (Masur 2006; Jones 2007, 2009).
Deception
Mimicry, so it has been argued, can at times fulfil some functions such as providing a way to deceive (e.g. pretending to be a predator or as a disguise of one’s own identity). In this context, it simply refers to mimicry of innocuous plants and animals to look like a poisonous, or at least dangerous, species so as to be left alone by predators. Such type of mimicry occurs in plants, invertebrates and even in vertebrates, but there is also another adaptation, exemplified by cuckoos, sneaking their own eggs into host nests.
In 2014, Herberstein and colleagues produced so much evidence of deception that they asked whether Australia is a hotspot for deception, using as their examples cuckoos, orchids and crabs. Australia happens to be rich in cuckoos, making up 20% of all cuckoo species in the world. Apparently, cuckoos are more successful in Australia than in Europe and America and the reasons, so they explain, may be related to the fact that in most Australian climate zones multi-nesting attempts are possible. Some orchids lure insects to pollinate them with sexual signals and as many as 50% of all deceptive orchids are found in Australia. Crab spiders lure pollinators to flowers with UV reflectors. The researchers argued that Australia’s milder climate and isolation fostered deception (Herberstein et al. 2014).
Indeed, some cuckoos employ mimicry as trickery (Davies 2011), whether as mimicked egg colour or in mimicry of host vocalisations. Great spotted cuckoos may mimic the host species (Redondo and Arias de Reyna 1988). Interestingly, common cuckoo chicks being raised by many different host species approximate the food begging calls of any of the host chicks by rhythm and frequency, despite the fact that they displace the host chicks by day 2 after hatching (Butchart et al. 2003).
Heterospecific mimicry is rare although one was reported of a South American species of greater racket-tailed drongos that issued a mimicked heterospecific alarm call of a nearby flock and thereby facilitated their escape from an aerial predator (Satischandra et al. 2010). The behaviour gets a little more complex when a bird has learned to give a false warning signal so that all other birds clear the area, allowing the caller to feed on the spoils. This behaviour of deception is then no longer mimicry but a type of a cognitively complex behaviour that will be discussed in more detail in Chapter 10 under ‘Deceptions and cheats’.
Mimicry can also represent a cheeky form of play behaviour or general social behaviour. The 75-year-old, permanently disabled galah that we adopted, for instance, learned to copy the calling of our dogs. He had learned the names of all four dogs and managed to mimic my voice so well that the dogs came running when he called. A consequence of this newly found power was that he regularly stood on the floor of the living room, called the dogs at least once a week, waiting until they had all assembled and then directed them to lie down. If anyone came too close to him, he opened his beak and, quite impressively, hissed at them. The dogs, incidentally, were Rhodesian ridgebacks – a very large breed – and to see a bird, barely standing high enough to reach a dog’s knee, pushing the dogs around and making them lie down was a sight to behold. This behaviour has also been noted in crows (J Marzluff pers. comm.) and has been observed in magpies. One example came from a family in rural New South Wales that had adopted a magpie and also had a cat and a dog. This magpie had learned to speak individual words, including, as in our galah, saying the name of their dog. The cat had made it amply clear that it wanted the magpie gone, which liked to sit provocatively, it seemed, in the middle of the yard on a tree stump. As the cat approached purposefully, the magpie did not move but called the name of the dog. The dog came running and chased the cat away. The magpie meanwhile preened itself as if in quiet triumph. Again, this did not just happen once but was played out with great regularity.
Mimicry of movement
It is now a widely held assumption that mirror neurons, at least in primates, play an important role in generating imitative movement behaviour. In 2005, Iacoboni and colleagues reported that mirror neurons can recognise the actions of others, by matching the observed action onto its motor counterpart encoded by the same neurons. Secondly, they found that it activated a neuronal chain of mirror neurons that can then encode the observed motor act in its entirety (i.e. encoding movement, and not something akin to still images). In 2008, Feenders and colleagues found the activation sites in songbirds, parrots and hummingbirds (all vocal learners). Importantly, vocal learning nuclei adjacent to discrete brain regions are also active during limb and body movements (Feenders et al. 2008). In this manner, an observed action is matched onto the observer’s motor repertoire (Iacoboni et al. 2005) leading to a tendency to automatically reproduce the action.
Brass and Heyes (2005) then teased apart the neural processes specifically related to the processing of observed actions, the selection of one’s own intended actions, the preparation for movement, and motor response execution. They concluded that motor imitation is based on the automatic activation of motor representations by movement observation. These externally triggered motor representations are then used to reproduce the observed behaviour. It was only a small step to conceive that species with the ability to perform vocal learning – as identified in humans, songbirds and some cetaceans – might also have developed ways of reading signals related to movement.
Birds had been considered incapable of motor imitation but in the last 15 years or so evidence of movement imitation has been produced in several species, such as pigeons (Klein and Zentall 2003; McGregor et al. 2006), Japanese quail (Dorrance and Zentall 2001; Akins et al. 2002), Carib grackles (Lefebvre et al. 1997), ravens (Izawa and Watanabe 2011) and European starlings (Fawcett et al. 2002). Such imitation is commonly defined as the ability of an observer to copy a feature or features of the body movement of a demonstrator.
There have been relatively few experiments assessing movement imitation by birds of the southern hemisphere. Only two species have so far been tested experimentally and both are psittacine species: the kea from New Zealand and the budgerigar.
Testing budgerigars on observational learning is not new (Dawson and Foss 1965; Galef et al. 1986). More recently, in 2002, Cecilia Heyes and Anna Saggerson tested imitative behaviour of the body movements of budgerigar juveniles to see whether they learned from observing demonstrators. The demonstrator was using its beak to remove one of two distinctively coloured objects (i.e. a blue and a black stopper) from a hole in the lid of a box and eating seed from within. Both objects could be removed by the lid either being pulled up or pushed down. They found that observer birds removed both stoppers in the same direction as their demonstrator and they tended to approach the same object as their demonstrator when the objects were distinguishable by both spatial and colour cues. They concluded that budgerigars are capable of motor imitation or emulation (Heyes and Saggerson 2002).
Several years later, in 2008 and 2009, respectively, Heyes and colleagues conducted another experiment using budgerigars (Mui et al. 2008; Richards et al. 2009). The experiment on a discrimination task was linked to a food reward. One budgerigar was trained either to peck for seeds on the floor (and then collect the reward) or put one foot on a small slightly raised button and that would release the food. Once the first bird was able to perform both ways of procuring food by either means, a second budgerigar was placed into an adjacent test box and then allowed to observe. The results showed that stepping demonstrators produced more stepping responses in observers and ground-pecking demonstrators produced more observers that pecked the ground. The researchers concluded that the ability to imitate depends on sensori-motor experience and phylogenetically ancient mechanisms of associative learning (Richards et al. 2009). In other words, mono-causal explanations tend not to work because there are multiple and complex interactions both within the brain and between the environment and the brain.
Cultural transmission via movement imitation
There is another aspect of the use of the beak for food collection or extraction that has caught the imagination of researchers for the past 100 years. It is the story of some British birds, especially of the blue tit. In 1921, in a town called Swaythling, birds were noted to tear bottle tops off milk bottles. Then the phenomenon spread and this was summarised in two papers by Hinde and Fisher (1951, 1972) in which they traced the spread of this hitherto unknown behaviour. It is now the most cited example of cultural transmission of a behaviour and it has also spawned a range of other theories, as well as opposition to the idea that the spreading of milk bottle opening represents the spread of culture (Lefebvre 1995).
Recently, a paper by Aplin and colleagues (Aplin et al. 2013) and a book by Bateson and Martin (2013) have offered fresh insight into the debate about the bottle top removal by these birds, adding concepts of innovation and creativity. When, as a natural part of its foraging skills, an avian species has learned to use the beak to tear off bark in order to get at insects below the bark, the step from that daily activity to tearing off bottle tops seems a small one because the motor patterns between the two activities may well be identical. However, this is misleading. First, as agreed by both Aplin and colleagues (2013) and Bateson and Martin (2013), the act of taking off tops of bottles is an innovative step, however small the step may appear to be. Second, it presupposes some learning by social facilitation or observation. It is successful as an innovation because there is evidence of its growing application in widening populations of the species. Moreover, Aplin and colleagues found significant age-and sex-biased social learning, with juvenile females being twice as likely to acquire the novel skill as birds of other age groups or males. Social learning was also more likely in subordinate males than in dominant males and that is noteworthy because both the readiness to learn and, possibly even to play, tends to be found in juveniles rather than in dominant (successful) adult individuals. Finally, as Bateson and Martin (2013) point out, at the basis of such innovations is also a certain level of creativity or exploration – why try to take a top off a milk bottle in the first place?
There is another element to the spread of the removal of milk bottle tops. Obviously, other birds must have watched the first demonstrator and then repeated the same action. Again, there are several ways to interpret this and it is possible to put the emphasis on different aspects of the copying act. Instead of an act performed mindlessly, it may be an act made possible because the brain of a bird, even one as small as that of a blue tit, is designed to deal with novelty and can incorporate the action into its repertoire.
In summary, imitation is said to represent a form of social learning that may require complex representations, not only from another’s perspective but also their intentions and how they relate to the individual performing the actions (see reviews in Whiten and Ham 1992; Heyes and Galef 1996). True imitation has been defined as ‘the copying of a novel or otherwise improbable act or utterance, or some act for which there is clearly no instinctive tendency’ (p. 135, Thorpe 1963) and as such the copied actions/vocalisations should not already be within the animal’s natural repertoire (Clayton 1978). In birds, two forms of imitative learning have been investigated: vocal mimicry and motor imitation.
Mirror neurons are thus not unique to the primate line but may be present in a wide range of birds. This means that mirror neurons have either been part of the brain for a very long time or they may have evolved independently a number of times.
Among the great advances in avian biology are three major milestones: the mapping of all song nuclei with all their complex feedback mechanisms and motor output routes, the complete mapping of the songbird genome; and, finally, the discovery and confirmation of the presence and function of mirror neurons in the avian brain. These parameters have created an entirely new framework for any assessment of learning ability and learning mechanisms in birds (Lefebvre 2013). The discovery of mirror neurons provides a biological basis for imitation that, in turn, can have explanatory power for variables generally held to be cognitively complex and related to theory of mind. This is not as far-fetched as it may seem.
Importantly, mimicry and imitation in birds, whether in the vocal or visuo-motor domain, has moved out of its spot of a relative oddity that was of little consequence and merely quaint. Along with the neuroscientific evidence of different kinds of forebrain activity, mimicry has taken its place as an important aspect of learning, of memory formation, and as a very important concomitant of group living, referred to as social learning.
Whatever reservation someone might have about imitation, there is no doubt about the ability of the brains of primates and birds to incorporate another’s movements and sounds into one’s own repertoire. In certain circumstances, on the basis of this ability perhaps it may be able to generate empathy (i.e. for a limping conspecific) or even lead to cooperation. That Heyes (2009), cited at the beginning of this chapter, perceived cooperation as one of the behaviours that mirror neurons may promote is very telling and relevant here because Australia boasts so many cooperative and pair bonded avian species. Remarkably also, Australia has a particularly high number of bird species that can do both: learn their vocalisations and mimic those of other species and even of other orders.