Jonathan M. Banksa; Kristelle Capistranoa; Pari Thakkara; Hemangi Ranadeb; Vaidik Sonia; Manali Dattab; Afsar R. Naqvia,* a Department of Periodontics, College of Dentistry, University of Illinois Chicago, Chicago, IL, United States
b Amity Institute of Biotechnology, Amity University Rajasthan, Jaipur, Rajasthan, India
* Corresponding author: email address: afsarraz@uic.edu
Abstract
Since the SARS-CoV-2 virus triggered the beginning of the COVID-19 pandemic, scientists, government officials, and healthcare professionals around the world recognized the need for accessible, affordable, and accurate testing to predict and contain the spread of COVID-19. In the months that followed, research teams designed, tested, and rolled out hundreds of diagnostic assays, each with different sampling methods, diagnostic technologies, and sensitivity levels. However, the contagious virus continued to spread; SARS-CoV-2 travelled through airborne particles and spread rapidly, despite the widening use of diagnostic assays. As the pandemic continued, hundreds of millions of people contracted COVID-19 and millions died worldwide. With so many infections, SARS-CoV-2 received many opportunities to replicate and mutate, and from these mutations emerged more contagious, deadly, and difficult-to-diagnose viral mutants. Each change to the viral genome presented potential added challenges to containing the virus, and as such, researchers have continued developing and improving testing methods to keep up with COVID-19. In this chapter, we examine several SARS-CoV-2 variants that have emerged during the pandemic. Additionally, we discuss a few major COVID-19 diagnostic technique categories, including those involving real-time PCR, serology, CRISPR, and electronic biosensors. Finally, we address SARS-CoV-2 variants and diagnostic assays in the age of COVID-19 vaccines.
Keywords
COVID-19; SARS-CoV-2; Diagnostics; Variants; RT-PCR; Serology; CRISPR; Biosensors; Sensitivity; Vaccines
Abbreviations
SARS-CoV-2 severe acute respiratory syndrome coronavirus 2
WHO World Health Organization
VOC variant of concern
B.1.1.7 SARS-CoV-2 alpha variant
B.1.351 SARS-CoV-2 beta variant
P.1 SARS-CoV-2 gamma variant
B.1.617.2 SARS-CoV-2 delta variant
E protein envelope protein
S protein spike protein
M protein membrane protein
N protein nucleocapsid protein
RBD receptor-binding domain
ACE2 angiotensin-converting enzyme 2
NTD N-terminal domain
RT-PCR real-time reverse transcription polymerase chain reaction
cDNA complementary DNA
USFDA United States Food & Drug Administration
EUA emergency use authorization
CDC United States Centers for Disease Control and Prevention
LOD limit of detection
SGTF spike-gene target failure
LFIA lateral flow immunoassay
ELISA enzyme-linked immunosorbent assay
CLIA chemiluminescent immunoassay
ECLIA electrochemiluminescence immunoassay
ECL electrochemiluminescence
CV2T dimension EXL SARS-CoV-2 total antibody assay
CRISPR clustered regularly interspaced short palindromic repeats
crRNA CRISPR-RNA
dsDNA double-stranded DNA
ssDNA single-stranded DNA
SHERLOCK specific high-sensitivity enzymatic reporter unlocking
DETECTR SARS-CoV-2 DNA endonuclease-targeted CRISPR Trans reporter
RT-LAMP loop-mediated amplification
qRT-PCR real-time quantitative reverse transcription polymerase chain reaction
SPR surface plasmon resonance
BAL bronchoalveolar lavage
IVD in vitro diagnostic
NAAT nucleic acid amplification test
UTR untranslated region
1: Introduction
In December 2019, the Severe Acute Respiratory Syndrome Coronavirus 2, better known as SARS-CoV-2, underwent zoonotic transmission to infect a human and cause a viral outbreak (Peñarrubia et al., 2020). On March 11, 2020, the World Health Organization (WHO) declared COVID-19 a pandemic (van Dorp et al., 2020). SARS-CoV-2 is genetically similar to a few other beta coronaviruses that infect animals, including bats (RaTG13) and pangolins (Salian et al., 2021). Believed to have originated due to mutation, SARS-CoV-2 spread from human to human and continued mutating as it rapidly divided in each host (van Dorp et al., 2020). With the world unable to contain SARS-CoV-2 or the surfacing mutants, the COVID-19 outbreak escalated to an epidemic and ultimately a pandemic, with high infection and death rates globally (Salian et al., 2021).
On November 12, 2021, the World Health Organization COVID-19 Dashboard reported 251,788,329 confirmed cases of COVID-19 as well as 5,077,907 deaths from the disease (WHO COVID-19 Dashboard, 2020). The virus spreads through air droplets, resulting in unique transmission patterns (Datta, Singh, & Naqvi, 2021). Countries worldwide experienced wave after wave of new cases, often followed by a chaotic spike in deaths (Dyer, 2021). To manage and prevent new COVID-19 waves, researchers must continue to study SARS-CoV-2 and develop COVID-19 diagnostic methods to contain the viral spread (Vandenberg, Martiny, Rochas, van Belkum, & Kozlakidis, 2020).
SARS-CoV-2 possesses a spherical shape of approximately 150 nm, and its genome is ~ 30 kb long single-stranded RNA (Datta et al., 2021). The viral genome was sequenced in its entirety and shared with the NCBI Genbank on January 5, 2020. In the following months, scientists identified thousands of additional sequences in countries around the world (Sallam & Mahafzah, 2021; van Dorp et al., 2020).
Since the onset of the pandemic, diagnostic assays have served as valuable tools in the fight against COVID-19. Many were developed as the first wave of viral cases surfaced. Most diagnostic assays helped detect viral infection in human samples or identify particular strains of SARS-CoV-2 through individual or community testing (Vandenberg et al., 2020). The need for both individual and community testing is clear: to monitor viral infection and prevent the spread of disease by isolating infected individuals. Further, identifying different viral mutants is necessary to monitor emerging dominant variants throughout the world and develop policies to counter their spread (Sallam & Mahafzah, 2021; Vandenberg et al., 2020). With that understanding, researchers continue to investigate key variants to better understand their differences in transmission and disease manifestation.
In a pandemic that has resulted in mask mandates, physical distancing, and rampant death and disease, diagnostic assays are a necessary component of global recovery efforts. To enter a post-pandemic world, the spread of COVID-19 must be sustainably contained. The end of the pandemic is only possible with the information that diagnostic assays provide. For people to return to their jobs, families, and lives safely, without the threat of undetected variants and uncontrollable infections, the development of effective and accessible diagnostics must be a worldwide priority.
With an ever-changing viral genome, diagnostic assays must continue to develop to keep up with new variants. Presently, diagnostics are used to identify viral infections in individuals and extrapolate viral prevalence information in broader populations with a more epidemiological approach (Uddin et al., 2020). Diagnostic assays have been used to survey sewage contents for viral prevalence, revealing trends in viral load throughout communities (Martin et al., 2020). These diagnostic strategies, coupled with individual testing and variant detection, provides a robust framework for the global diagnostic arsenal needed to combat the viral spread.
Regarding individual testing techniques, there are many. They range in specificity, sensitivity, and accuracy, and they each have their benefits and drawbacks. In this chapter, we focus on the following diagnostic methods: real-time polymerase chain reaction, serology, CRISPR, and electronic biosensors. We will discuss the strengths and limitations of viral detection techniques, and their role in diagnosing SARS-CoV-2 variants.
2: SARS-CoV-2 variants of concern
The rapid spread of SARS-CoV-2 around the globe and its consequential high viral replication rates have increased the chance of mutation events. As a result, numerous genetic variants have emerged since the discovery of the original strain (Wuhan-Hu-1) in December 2019 (Shahhosseini, Babuadze, Wong, & Kobinger, 2021). As of October 2021, there were four COVID-19 strains classified as “variants of concern” (VOCs): (1) B.1.1.7 (Alpha) (2) B.1.351 (Beta) (3) P.1 (Gamma) and (4) B.1.617.2 (Delta) (Table 1). These variants display key mutations, which mounting evidence has shown to confer one or more of the following viral attributes: increased transmissibility, increased disease severity, decreased neutralizing antibody response acquired through natural infection or vaccination, decreased treatment/vaccine efficacy, and diagnostic detection failures.
Table 1
|
Notable variations of the variants of concern (VOCs). Important mutations are notated in bold. |
||
|
Variant |
Mutation |
Gene |
|
B.1.1.7 (Alpha) |
TI001I |
ORF1a/b |
|
L18F (some isolates) |
S |
|
|
Q27 stop |
ORF8a/b |
|
|
D3L |
N |
|
|
B.1.351 (Beta) |
K1655N |
ORF1a/b |
|
L18F (some isolates) |
S |
|
|
P71L |
E |
|
|
T205I |
N |
|
|
P.1 (Gamma) |
S1188L |
ORF1a/b |
|
L18F |
S |
|
|
E92K |
ORF8a/b |
|
|
P80R |
N |
|
|
B.1.617.2 (Delta) |
P323L |
ORF1a/b |
|
T19R |
S |
|
|
S26L |
ORF3a |
|
|
I82T |
M |
|
|
V82A |
ORF7a |
|
|
119–120 del |
ORF8a/b |
|
|
R203M |
N |
|
Although mutations have accumulated throughout SARS-CoV-2 genes encoding the ORF1ab, ORF3, ORF8, and N (nucleocapsid) proteins, mutations in the S (spike)-protein are particularly significant. The S-protein is a type I transmembrane protein that initially exists as an inactive precursor. During the early stages of infection, SARS-CoV-2 uses its S-protein's receptor-binding domain (RBD) to engage the angiotensin-converting enzyme 2 (ACE2) host receptor, which is primarily expressed in lung and intestinal cells. Following the RBD-ACE2 binding, proteolysis occurs at the S1/S2 junction by furin or other proteases (i.e. TMPRSS2 or cathepsin proteases), exposing a second cleavage site within the S2, subsequently liberating the S2 fusion peptide to initiate the viral-host membrane fusion and leading to S1 shedding (Peacock et al., 2021). Specific mutations such as D614G result in less S1 shedding and increased incorporation of the S protein into the virion, thereby enhancing viral infectivity.
Because the S-protein plays a crucial role in viral invasion and host cell attachment, it is a major target for antibody therapy, specifically neutralizing antibodies that target SARS-CoV-2. Within the S protein, the RBD and the NTD (N-terminal domain) are primary targets. NTD-targeting and RBD-targeting antibodies form complexes with the NTD and RBD, respectively, preventing viral entry and host cell-virus membrane fusion. However, some variants such as B.1.351 (Beta) and P.1 (Gamma) have developed mutations that prevent or weaken the binding of particular antibodies to the S-protein (Yang & Du, 2021). This section highlights changes in the viral genome, spread, and detection of each VOC.
2.1: B.1.1.7 (Alpha) variant
In September 202, the two earliest documented genomic samples of lineage B.1.1.7 (also known as Alpha variant, 20I/501Y·V1, or VOC 2020 12/01) were collected in Kent and Greater London. The variant became notable for its unusually large number of genomic changes, accruing 14 lineage-specific amino acid substitutions before its detection, and it quickly became predominant worldwide (Morris et al., 2021). By February 2021, 90 countries/territories reported cases of B.1.1.7 (Alpha) (Shahhosseini et al., 2021).
In total, the B.1.1.7 (Alpha) variant has 23 mutations (17 amino acid changes) from the Wuhan-Hu-1 strain (Fig. 1) (Abdool Karim & de Oliveira, 2021). Notable mutations of B.1.1.7 (Alpha) include the spike D614G mutation, spike N501Y mutation, spike HV 69–70 deletion, and other mutations that are not addressed in this text. B.1.1.7 (Alpha) features a significant number of non-synonymous amino acid substitutions, which multiple sources implicate for its increased transmissibility and infectivity; experts estimate that B.1.1.7 (Alpha) is up to 70% more transmissible than the Wuhan-Hu-1 strain (Cheng et al., 2021; Jackson, Zhang, Farzan, & Choe, 2021; Meng et al., 2021). Additionally, B.1.1.7 (Alpha) appears to be resistant to neutralization by most NTD-directed and a few RBD-directed monoclonal antibodies (but not convalescent plasma or vaccine sera) due to the N501Y mutation (Wang et al., 2021).
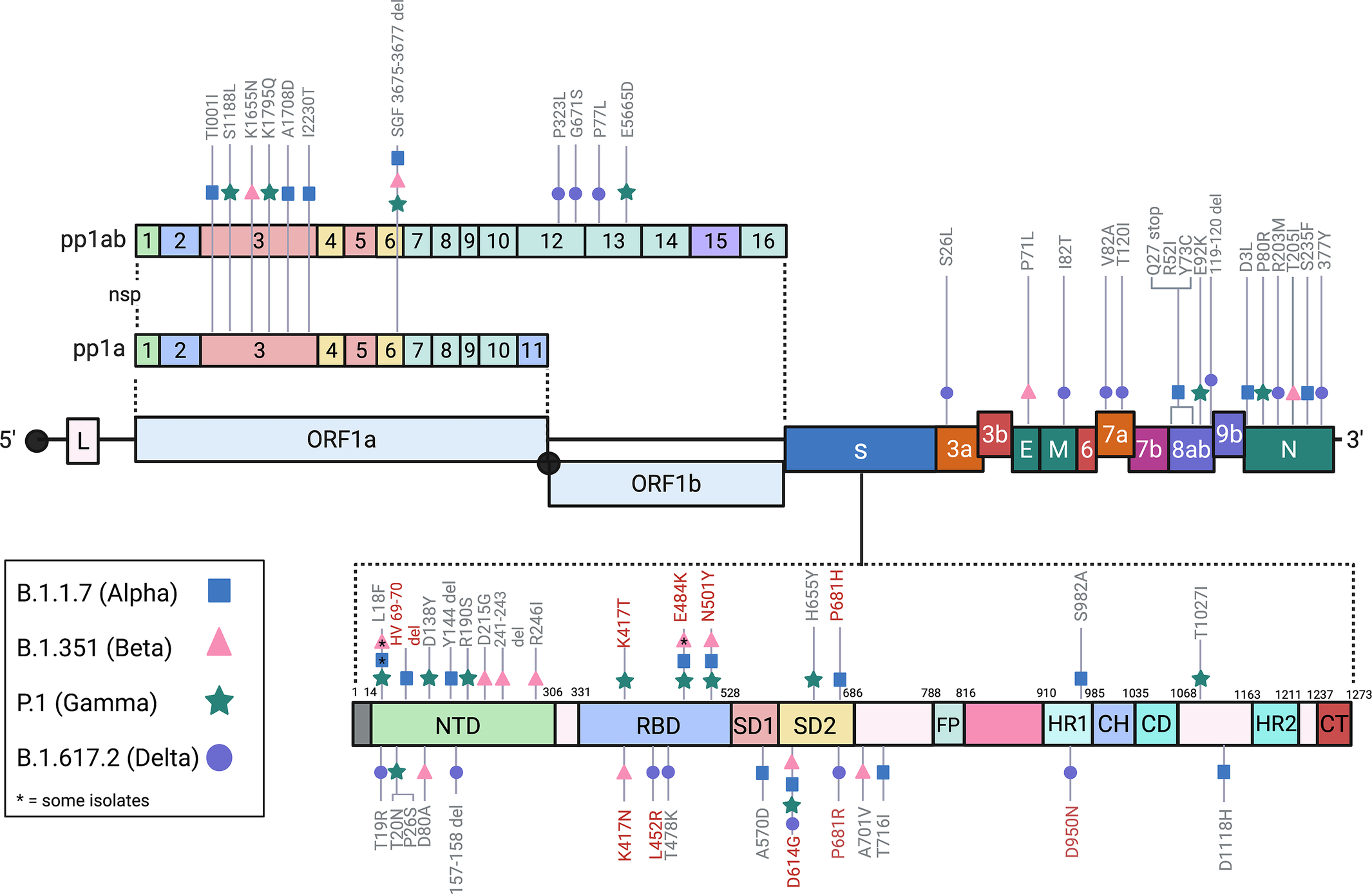 Fig. 1 SARS-CoV-2 genome and the notable mutations of the Variants of Concern (VOC). Crucial mutations notated in red (BioRender.com, 2021).
Fig. 1 SARS-CoV-2 genome and the notable mutations of the Variants of Concern (VOC). Crucial mutations notated in red (BioRender.com, 2021).
2.2: B.1.351 (Beta) variant
In October 2020, the second wave of COVID-19 infections swarmed through Nelson Mandela Bay in South Africa and spread through the Western Cape, Eastern Cape, and KwaZulu-Natal provinces within weeks. At the peak of the second wave, daily confirmed cases of COVID-19 in the area rose as high as > 20% of PCR tests at the local municipality level. Most of these cases were later attributed to a new variant, B.1.351 (Beta). The B.1.351 (Beta) variant emerged independently of B.1.1.7 (Alpha), likely after the first wave of SARS-CoV-2 cases in the Eastern Cape province (Tegally et al., 2021). B.1.351 (Beta) contains multiple mutations in the S-protein, three of which are in the RBD (N501Y, E484K, and K417N) and alter the binding affinity for ACE2 receptors (Fig. 1).
Like the B.1.1.7 (Alpha) variant, B.1.351 (Beta) also increases the risk of transmission. To a greater degree compared to B.1.1.7 (Alpha), however, there are multiple reports that the B.1.351 (Beta) reduces neutralization by monoclonal antibodies (against the N-terminal and RBD domains), convalescent plasma (9.4-fold), and post-vaccination sera (10.3- to 12.4-fold) compared to wild type SARS-CoV-2 (Wang et al., 2021). The increased transmissibility of B.1.351 (Beta) is likely due to mutations D614G and N501Y, which also contributed to increased transmissibility in the B.1.1.7 (Alpha) variant. In B.1.351 (Beta), the spike E484K and K417N mutations also contribute to immune escape by enabling the evasion of antibody and plasma neutralization (Greaney et al., 2021; Harvey et al., 2021; Laffeber, de Koning, Kanaar, & Lebbink, 2021; Wang et al., 2021).
2.3: P.1 (Gamma) variant
The P.1 (Gamma) variant (alias of B.1.1.28.1) was first reported in January 2021 when four travellers arrived at a Japanese airport from Brazil. Molecular clock analysis shows that P.1 (Gamma) emerged around November 2020 and was responsible for a resurgence of cases in Manaus, the capital of the Amazonas. In late December 2020, a reported 42% of specimens from Manaus tested positive for the P.1 (Gamma) variant. The spike of cases was alarming because approximately 75% of Manaus residents have previously tested positive for SARS-CoV-2 and were expected to have a degree of immunity (Faria et al., 2021). Today, there are approximately 62 countries with the P.1 (Gamma) sequence. In total, P.1 (Gamma) has 17 mutations (11 amino acid changes).
Although distinct, P.1 (Gamma) and B.1.351 (Beta) share several characteristics. First, P.1 (Gamma) and B.1.351 (Beta) feature the same key spike mutations N501Y and E484K in the RBD of the S-protein. While P.1 (Gamma) and B.1.351 (Beta) have acquired K417T and K417N, respectively, both mutations are highly similar in effect and are often grouped. Similar to B.1.351 (Beta), N501Y is likely responsible for P.1 (Gamma)’s increased binding to the ACE2 receptor and consequential increased transmissibility. Moreover, E484K and K417T also reportedly result in a dampened neutralizing humoral immunity response. Despite these similar RBD mutations, Dejnirattisai et al. suggest that natural and vaccine-induced antibody neutralization are significantly more effective against P.1 (Gamma) than B.1.351 (Beta); the mechanism behind this difference in antibody neutralization of P.1 (Gamma) and B.1.351 (Beta) is unclear. Nonetheless, the fact that P.1 (Gamma) and B.1.351 (Beta) independently evolved similar RBD mutations indicates convergent evolution (Dejnirattisai et al., 2021).
2.4: B.1.617.2 (Delta) variant
B.1.617.2 (Delta) is a subtype of the SARS-CoV-2 B.1.617 lineage, which first emerged in October 2020 in India. Compared to the other B.1.617 subtypes, B.1.617.1 (Kappa) and B.1.617.3, B.1.617.2 (Delta) is considered the most transmissible. As of August 2021, B.1.617.2 (Delta) spread to at least 124 countries and serves as a significant cause of concern due to its increased transmissibility relative to all other SARS-CoV-2 strains. Current estimates show that B.1.617.2 (Delta) is 60% more contagious than B.1.1.7 (Alpha). Interestingly, some data indicate that the B.1.617.2 (Delta) variant results in a shorter incubation period than the original strain (B.1.617.2 (Delta): mean of 4 days; Wuhan-Hu-1: mean of 6 days). Moreover, the high viral load in people infected with the variant (up to 1260 times higher than those infected with the original strain) further heightens the transmissibility of B.1.617.2 (Delta) and could play a part in breakthrough infections (Li et al., 2021; Reardon, 2021). Finally, one study found that B.1.617.2 (Delta) remains infectious for longer (B.1.617.2 (Delta): 18 days; Wuhan-Hu-1: 13 days) (Ong et al., 2021). Altogether, these data indicate that B.1.617.2 (Delta) replicates faster and at higher rates compared to other mutants.
There are 12 mutations in the B.1.617.2 (Delta) genome, 10 of which are in the S-protein: T19R, G142D (in some strains), 156del, 157del, R158G, L452R, T478K, D614G, P681R, and D950N (Fig. 1). L452R and T478K both confer a significant increase in immune evasion and infectivity to the B.1.617.2 (Delta) variant, and they interfere with the host antibody response (monoclonal antibodies and sera from convalescent and vaccinated individuals) by weakening the binding between antibodies and the S-protein (Di Giacomo, Mercatelli, Rakhimov, & Giorgi, 2021; Motozono et al., 2021; Planas et al., 2021). P681R is located in the S-protein S1/S2 (furin) cleavage site, and it is another mutation that confers better transmissibility (Cherian et al., 2021).
The VOCs share key mutations, mainly in the RBD and NBD-containing S1 subunit. Consequently, a significant portion of these major spike mutations increases the ACE2 binding affinity, altering viral pathogenicity and virulence. Not to mention, specific mutations appear to have compensatory effects when combined. One example is the K417N: N501Y combination seen in B.1.351 (Beta). K417N reportedly reduces ACE2 binding affinity by disrupting the formation of two salt bridges in the RBD-ACE2 complex. Despite K417N, B.1.351 (Beta) is still estimated to be 2.5 times more transmissible than Wuhan-Hu-1 due to the N501Y mutation (Harvey et al., 2021).
The effect of these mutations on vaccine efficacy sensitivity is of great concern. While SARS-CoV-2 vaccines mRNA-1273 (Moderna) and BNT162b2/Comirnaty (Pfizer/BioNTech) reportedly elicit a robust antibody response against B.1.1.7 (Alpha) and P.1 (Gamma), the neutralization effect is significantly weaker against B.1.351 (Beta). Data regarding the effect of B.1.617.2 (Delta) on the vaccines is limited. However, early reports suggest that while protection against infection is dampened, mRNA-1273 and BNT162b2 still vigorously protect against hospitalization and death (Khateeb, Li, & Zhang, 2021).
3: COVID-19 diagnostics
A prevalent concern is the effect of mutations on diagnostic sensitivity. With the background provided regarding the SARS-CoV-2 virus and its variants, the diagnostic field is rapidly evolving. In this text, we discuss the various widely used diagnostic strategies that researchers developed to test for the virus and its effects on the body (as manifested through antibody response). The sections below describe four SARS-CoV-2 diagnostics approaches: RT-PCR, Serology, CRISPR and Biosensor/electric/smart sensors (Fig. 2). Additionally, we discuss how diagnostic techniques have been designed and redesigned for the detection of SARS-CoV-2.
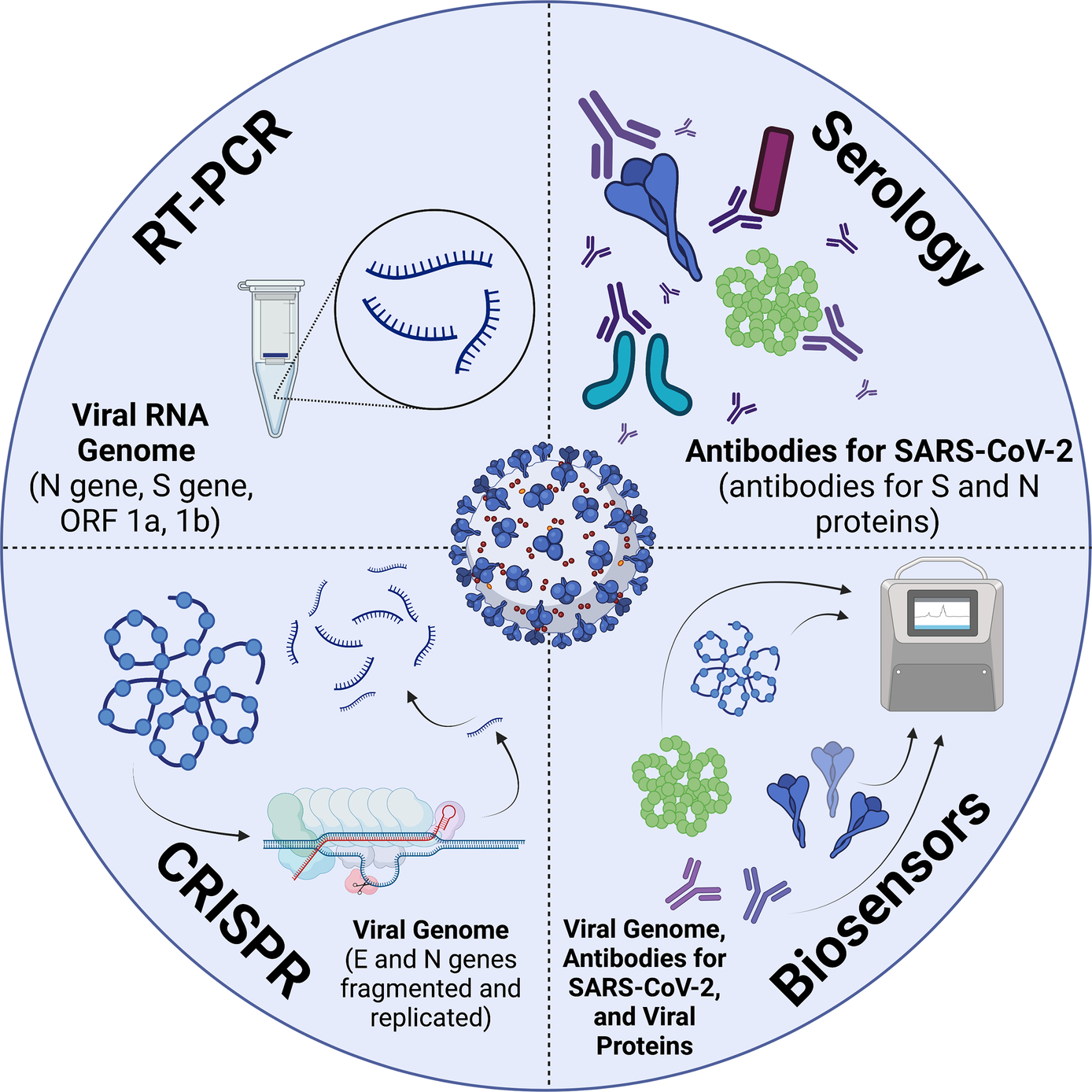 Fig. 2 COVID-19 Diagnostic Techniques and their Targets (BioRender.com, 2021).
Fig. 2 COVID-19 Diagnostic Techniques and their Targets (BioRender.com, 2021).
3.1: Real-time PCR diagnostics
Real-time reverse transcription polymerase chain reaction (RT-PCR) is a widely known technique involving a series of chemical reactions with several significant applications related to genetics. For decades, polymerase chain reactions have been used to selectively amplify target genetic molecules, and PCR protocols have since been adapted to utilize a multitude of sample types (Nolan, Hands, & Bustin, 2006). The integration of a fluorescent indicator allowed for relative molecular quantities to be observed and compared in real time (Heid, Stevens, Livak, & Williams, 1996; Morley, 2014). Since then, RT-PCR methods have allowed researchers to quantify and amplify expressed RNA (Green & Sambrook, 2018), study genetic expression on the microscale in microdissected cells (Paweletz, Charboneau, & Liotta, 2001), and, in conjunction with electrospray ionization mass spectrometry (ESI/MS), perform viral assays and diagnose pathogens (Deyde, Sampath, & Gubareva, 2011; Wolk, Kaleta, & Wysocki, 2012). As with other pathogen diagnostics, RT-PCR tests play a significant role in providing public health experts and government officials with valuable information regarding viral presence and spread (Farasani, 2021).
At its core, RT-PCR involves the following processes: RNA is reverse transcribed into complementary DNA (cDNA), then the cDNA is replicated exponentially with each experimental cycle, and finally the expression levels of the cDNA are observed in real time by fluorescent indicators as a proxy, and the expression levels are used to extrapolate information regarding the genetic expression and activity within a sample (Nolan et al., 2006). These samples can come from a variety of sources, as previously mentioned, and PCR tests can probe for the presence of several known pathogens (Fig. 3).
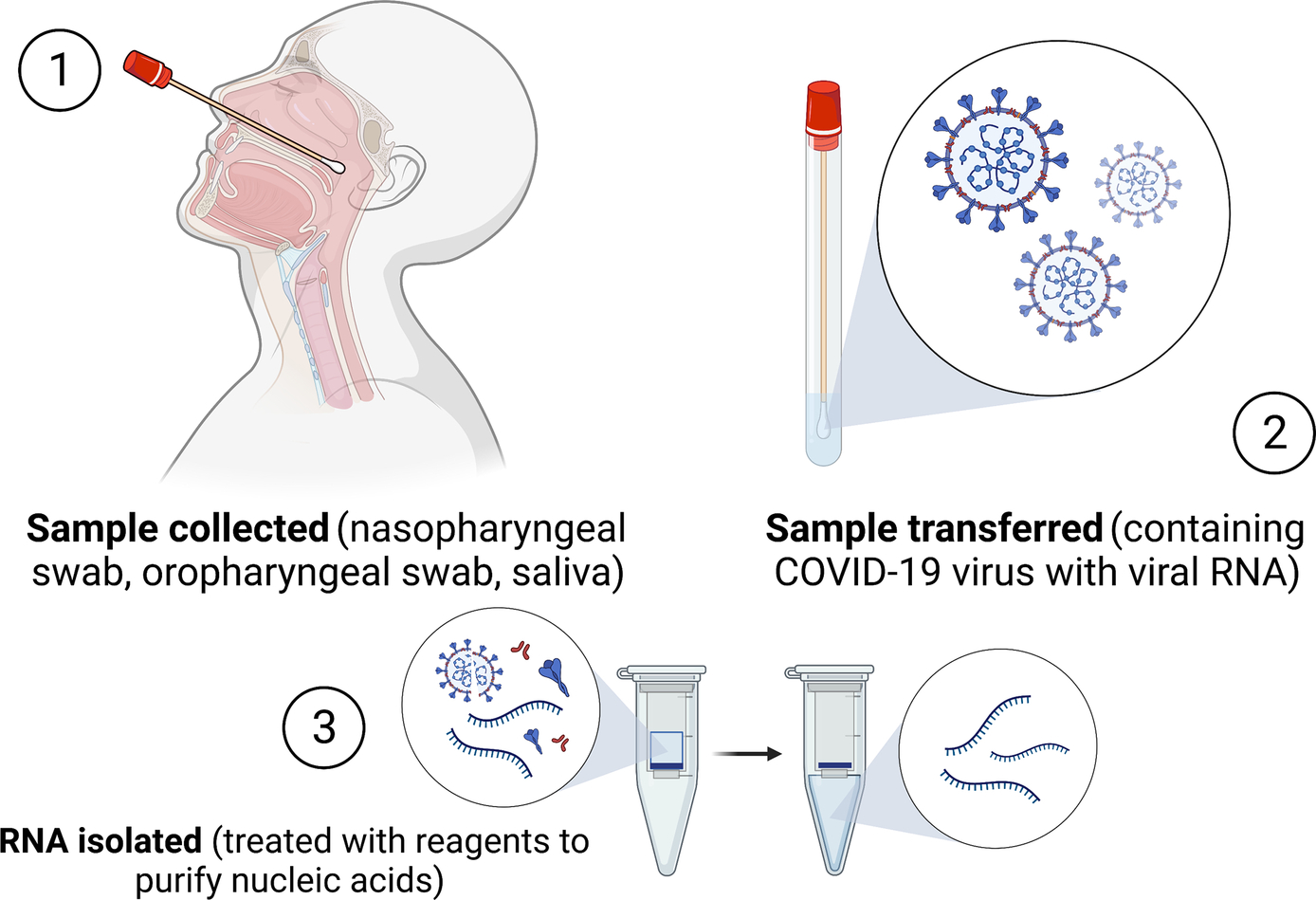 Fig. 3 RT-PCR Diagnostic Process. Schematic showing the description of obtaining a sample, isolating RNA and RT-qPCR using instrument to detect viral genome (BioRender.com, 2021).
Fig. 3 RT-PCR Diagnostic Process. Schematic showing the description of obtaining a sample, isolating RNA and RT-qPCR using instrument to detect viral genome (BioRender.com, 2021).
3.1.1: RT-PCR and COVID-19 detection
With potential applications ranging from forensics to genomics and microbiology, RT-PCR has also emerged as a valuable viral diagnostic assay, especially during the COVID-19 pandemic (Bustin, Benes, Nolan, & Pfaffl, 2005). Perhaps the most esteemed method of PCR diagnostics, and SARS-CoV-2 diagnostics at large, involves the isolation and purification of nucleic acids before the PCR and reverse transcription occurs. This method is praised for its accuracy, specificity, and sensitivity, all of which make it a popular choice for diagnosing SARS-CoV-2 worldwide (Fomsgaard & Rosenstierne, 2020). The accuracy of commercial RT-PCR tests for COVID-19 were experimentally validated, and the tests were found to meet international standards for accuracy (Wu, Xu, Zhu, & Xia, 2020). In addition to detecting viral presence in samples, RT-PCR tests have yielded valuable information about viral load and the infectivity of different COVID-19 variants (Korber et al., 2020). Though they are not considered true rapid tests, in comparison to other methods used for measuring RNA expression, PCR reactions are faster and safer, requiring fewer dangerous materials for the tests (Farasani, 2021; Green & Sambrook, 2018).
There are several RT-PCR assays that governing bodies have approved for use in testing for COVID-19. To rapidly respond to the developing COVID-19 pandemic, the United States Food & Drug Administration (USFDA) issued Emergency Use Authorizations (EUA) to quickly authorize the use of RT-PCR diagnostics. According to the USFDA, 210 RT-PCR diagnostic assays have been granted such approval at the time of writing (https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas-molecular-diagnostic-tests-sars-cov-2). Two such tests were developed by the United States Centers for Disease Control and Prevention (CDC), and the tests fall into two separate categories of RT-PCR diagnostic assays: simplex and multiplex assays (CDC, 2021). As detailed in Table 2, both tests target the nucleocapsid (N) protein-coding gene in SARS-CoV-2. However, the multiplex assay also tests for genes in the Influenza A and B viruses, allowing it to test for co-infection of the viruses that manifest themselves similarly to COVID-19 (Shu et al., 2021). Table 2 depicts their sensitivities as both > 95%. The TaqPath COVID-19 Combo Kit by Thermo Fisher tests for open reading frames 1a and 1b, the spike protein-coding region, and the N protein-coding region of the SARS-CoV-2 genome (www.thermofisher.com/us/en/home/clinical/clinical-genomics/pathogen-detection-solutions/covid-19-sars-cov-2/multiplex.html; https://www.fda.gov/media/136112/download). With a 100% detection rate at the limit of detection (LOD), this highly sensitive test can detect various viral mutants because of its multi-site screening capabilities. Similarly, Labcorp's COVID-19 RT-PCR Test is able to screen for multiple parts of the viral genome, resulting in a high sensitivity of > 95% at the LOD (https://www.fda.gov/media/136151/download). However, the invasive nature of the nasal or bronchial swab required for these assays can cause subject discomfort.
Table 2
|
RT-PCR diagnostic assays and their gene targets, sensitivities, limits of detection, and other key details. |
||||
|
Type of test, company, source |
Approval and date |
Targets |
Sensitivity and limit of detection (LOD) |
Pros and cons |
|
Influenza SARS-CoV-2 Multiplex Assay (Multiplex), CDC (CDC, 2021; https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas-molecular-diagnostic-tests-sars-cov-2; Shu et al., 2021) |
USFDA EUA, 07/02/2020 |
SARS-CoV-2 N gene, Influenza A Matrix (M1) gene, Influenza B nonstructural 2 (NS2) gene |
greater than 95% detection rate at LOD |
Pros: Tests multiple viruses, high sensitivity |
|
2019-nCoV RT-PCR Diagnostic Panel – QIAGEN EZ1 Advanced XL (Singleplex), CDC (CDC, 2021; https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas-molecular-diagnostic-tests-sars-cov-2) |
USFDA EUA, 02/04/2020 |
SARS-CoV-2 N gene |
greater than 95% detection with LOD 100.5 RNA copies/μL |
Pros: High sensitivity and accuracy |
|
TaqPath COVID-19 Combo Kit (Singleplex), Thermo Fisher Scientific, (https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas-molecular-diagnostic-tests-sars-cov-2; www.thermofisher.com/us/en/home/clinical/clinical-genomics/pathogen-detection-solutions/covid-19-sars-cov-2/multiplex.html; https://www.fda.gov/media/136112/download) |
USFDA EUA, 03/13/2020 |
SARS-CoV-2 ORF 1a, ORF 1b, S, and N genes |
100% detection rate at LOD |
Pros: Tests multiple genes, differentiates variants |
|
COVID-19 RT-PCR Test (Singleplex or Multiplex), Laboratory Corporation of America (Labcorp) (https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas-molecular-diagnostic-tests-sars-cov-2; https://www.fda.gov/media/136151/download) |
USFDA EUA, 03/16/2020 |
SARS-CoV-2 nucleocapsid (N) gene |
greater than 95% detection rate at LOD |
Pros: Targets multiple genes, versatile |
|
SalivaDirect (Dualplex), Yale School of Public Health (https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas-molecular-diagnostic-tests-sars-cov-2; https://ysph.yale.edu/salivadirect/publications/Vogels et al., 2021) |
USFDA EUA, 08/27/2021 |
SARS-CoV-2 nucleocapsid (N) gene |
94% positive agreement when compared to TaqPath COVID-19 combo kit, no LOD specified |
Pros: Efficient, noninvasive swab sample |
Lastly, the most recent assay to be granted EUA approval at the time of writing came out of the Yale School of Public Health, the SalivaDirect dualplex RT-PCR assay. This assay yields comparable sensitivity of 94% agreement when compared to the TaqPath assay, and its less invasive salivary sample requirement provides a significant advantage without completely sacrificing sensitivity (https://ysph.yale.edu/salivadirect/publications/; Vogels et al., 2021). These assays are described in more detail in Table 2. Alternative RT-PCR techniques with fewer required reagents were developed in response to a shortage in key RT-PCR reagents, and articles by Fomsgaard and Rosenstierne (2020) and Smyrlaki et al. (2020) provide a more comprehensive description of these alternate approaches.
3.1.2: RT-PCR and COVID-19 variants
Despite the many benefits of RT-PCR in diagnostics, the testing method is not without its flaws, especially when it comes to the variants of COVID-19 and false negative test results. If a PCR test fails to detect a significant amount of viral RNA in a sample, resulting in a negative test result, and another test later reveals a positive test result for the virus, the original test is deemed a “false negative” (Arevalo-Rodriguez et al., 2020). These inaccuracies in testing have significant consequences, allowing unknowing individuals to potentially spread a virus while believing they are uninfected. A review article discovered a high propensity for false negative PCR results in patients with COVID-19 symptoms, and authors advocated for multiple tests if a patient is suspected of having a false negative (Arevalo-Rodriguez et al., 2020).
Even when multiple tests are administered, false negatives still impact PCR testing results. A case study featured a 63-year old female with symptoms of COVID-19 who repeatedly tested negative for the virus via a nasopharyngeal swab (Shukha, Makhoul, Abu-Elhija, Hayek, & Hamoud, 2021). Upon testing her for COVID-19 using a sample acquired during a bronchoscopy, the results came back positive for COVID-19, and those results were followed by even more negative results from the nasopharyngeal RT-PCR diagnostic. Thus, due to different viral manifestation patterns, as evidenced by the RT-PCR test results, the standard nasopharyngeal swab was an inaccurate testing diagnostic for the patient already displaying COVID-19 symptoms.
Differences in viral manifestation and behaviour are often attributed to viral mutations, forming variants of COVID-19 with different genome sequence, transmissibility and clinical manifestations. Some variants can evade RT-PCR detection because of mutations in the target areas of RNA reverse transcription that prevent gene amplification. Other variants feature mutants that cause conformational changes in proteins that influence gene accessibility, rendering RT-PCR assays ineffective. For example, the H69-V70 deletion of the B.1.1.7 (Alpha) variant impacts the accuracy of diagnostic tests that target the S-gene, as is the case for the ThermoFisher TaqPath real-time PCR diagnostic kit. Specifically, the mutation likely induces a conformational change in the S protein, which interferes with the binding of the primers of Taqpath and other similar PCR tests to the S-gene target, resulting in “S-gene target failure” (SGTF) (Borges et al., 2021). SGTF is a phenomenon in which RT-PCR testing fails to amplify and thereby detect the S gene in an otherwise positive PCR test.
These challenges and others require innovative solutions to monitor the ever-changing variants of COVID-19. In the case of variant-induced spike-gene target failure, areas with high B.1.1.7 (Alpha) prevalence, such as England and Portugal, have used SGTF as a proxy to monitor the geographical dispersion and frequency of the variant (Borges et al., 2021). One study found a high correlation between B.1.1.7 (Alpha) frequency and SGTF in community-based diagnostic PCR testing in the UK, allowing the researchers to distinguish between VOC and non-VOC incidence by region over time (Volz et al., 2021). Due to the multiple gene targets of the ThermoFisher TaqPath assay, including ORF1 and N genes, the test remains a functionally reliable detector of SARS-CoV-2 overall, as noted in Table 2. It is worth noting that many commercially available SARS-CoV-2 diagnostic tests have multiple genetic targets and do not use the S-gene as the primary target, including multi-target RT-PCR tests in Table 2. Currently, there is no evidence of S-gene mutations in other variants of concern and mutations in other genes affecting the accuracy of diagnostic tests (Babb de Villiers, Blackburn, Cook, & Janus, 2021). All-in-all, diagnostic tests are expected to remain accurate despite further viral mutations.
Similar improvements in other diagnostic assays have led to continued effectiveness in RT-PCR testing. In one study, RT-PCR tests that targeted multiple genetic sites were able to maintain a high sensitivity level despite the mutations that would typically decrease sensitivity in a RT-PCR test with only one target region (Peñarrubia et al., 2020). A similar technique was used in England to monitor community viral presence through analysing sewage contents. By taking samples from the sewers and testing them with multi-target RT-PCR diagnostics, researchers were able to discover the presence and prevalence of different COVID-19 variants based on the target gene amplification and the expression levels (Martin et al., 2020). The results from this study verified the efficacy of lockdown measures in England, as the community viral load decreased during the enforced physical distance, and the findings also showcased the ability of RT-PCR diagnostics to test and influence health guidelines on an environmental scale.
Though there remain limitations to its efficacy, the benefits of RT-PCR presently outweigh the drawbacks as a diagnostic tool for COVID-19. It is used worldwide because it provides a sensitive, low cost, and safe option for viral diagnostics (Farasani, 2021). Additionally, despite the shortage of reagents involved in the RNA isolation process, techniques like heat treatment and lysis buffers provide alternatives that remain scalable, affordable, and safe (Fomsgaard & Rosenstierne, 2020; Smyrlaki et al., 2020). While variants have coincided with notable false negative results, developments in multi-target RT-PCR testing have produced high sensitivity tests that can distinguish between different variants (Martin et al., 2020). Although RT-PCR is not the rapid diagnostic tool, its effectiveness and timeliness makes it a useful technique for diagnosing en masse.
3.2: Serology based COVID-19 diagnostics
Serologic (antibody) tests are blood tests that measure the extent to which an individual has developed antibodies against different parts of a certain virus (https://www.healthline.com/health/serology; https://publichealth.jhu.edu/2021/variants-vaccines-and-what-they-mean-for-covid-19-testing). Antigens (e.g. bacteria, fungi, viruses, parasites) are substances that cause an immune response in the host, and specific antibodies are produced by the immune system to defend against these antigens (https://www.healthline.com/health/serology). Thus, the measurement of antibodies will provide further insight on the host's potential to fight viral infection and may even provide some insight into the virus pathobiology (Datta et al., 2021). Additionally, serology assays are more important in cases that do not allow for RNA isolation, either due to difficulty or no longer being present at the target site, as well as for epidemiological and post-vaccination monitoring studies (Zhang et al., 2020). Serological tests can also aid with autoimmune disorder diagnosis, in the case of the body mistaking its own healthy tissues for foreign invaders thus resulting in unnecessary antibody production.
There are several types of serological tests, including: rapid diagnostic tests, enzyme-linked immunosorbent assays, chemiluminescent immunoassays, electrochemiluminescence immunoassays, lateral flow chromatographic immunoassays and neutralization assays (Datta et al., 2021). Rapid serology diagnostic tests (e.g. lateral flow immunoassay [LFIA]) are convenient to implement at point-of-care, rapid and inexpensive, thus allowing for commercial kit production (Yamamoto et al., 2021). However, there is still a considerable amount of uncertainty with accuracy of rapid serologic tests.
Enzyme-linked immunosorbent assays (ELISA) rely on antigen binding to antibodies in a laboratory (https://www.healthline.com/health/elisa). Chemiluminescent immunoassays (CLIA) are also laboratory-based and rely on enzyme labelled antibodies and antigens (https://www.lornelabs.com/news-events/blog/what-is-chemiluminescent-immunoassay). One advantage of this assay technique is that its sensitivity levels are very high. Electrochemiluminescence immunoassays (ECLIA) rely on quantifying antigen or antibody, based on the change in electrochemiluminescence (ECL) signal, both before and after immunoreaction. ECLIA has increased sensitivity and can be more selective than other methods (Wu & Ju, 2012). Lateral flow chromatographic immunoassays rely on the collected specimen's IgM or IgG antibodies binding to labelled SARS-CoV-2 antigens to form a complex, which will later be accounted for (https://www.fda.gov/media/144071/download). Neutralization assays measure the levels of neutralizing antibodies from either recent or prior infection, which help inform the reasoning behind existing antibodies (https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-first-test-detects-neutralizing-antibodies-recent-or). Due to lower sensitivity of serological tests compared to molecular methods, serology tests are primarily used for retrospective diagnosis (Datta et al., 2021).
SARS-CoV-2 has four structural surface proteins: E, S, M and N proteins (envelope, spike, membrane, and nucleocapsid, respectively) (Datta et al., 2021). Serology tests against SARS-CoV-2 have been created by several companies and have been given emergency-use approval (EUA) via the U.S. Food and Drug Administration (FDA) (Zhang et al., 2020). Other countries that have also similarly approved serological tests include Germany, Singapore, China, Japan and Spain. Most serological tests have been developed to detect the presence of IgM/IgG antibodies against N, S and Receptor-Binding Domain (RBD) proteins in human plasma and serum (Ishikawa et al., 2009). Therefore, each serological test may be used for a single or a few variants, until a new variant emerges with mutations in the protein targeted by the serology test (https://publichealth.jhu.edu/2021/variants-vaccines-and-what-they-mean-for-covid-19-testing). As a result, serology diagnostics should not be used for vaccine efficacy testing, due to different vaccine targets. However, with further testing, serological diagnostic tests for COVID-19 may be used to inform a level of immunity that could perhaps decrease the severity of re-infection and the time during which the infection remains.
There are several serology diagnostic assays that are currently being studied for use against COVID-19 and its several variants (Table 3). FDA authorizes each for use under the Emergency Use Authorization (EUA). The Elecsys AntiSARS-CoV-2 S test uses an electrochemiluminescence quantitative immunoassay, and its overall sensitivity is 99.5%, post 14 days of PCR confirmation (https://www.centerforhealthsecurity.org/covid-19TestingToolkit/testing-basics/types-of-COVID-19-tests/serology-tests.html). SARS-CoV-2 antibodies against the spike RBD (Receptor Binding Domain) of the spike (S) protein (https://diagnostics.roche.com/us/en/products/params/elecsys-anti-sars-cov-2.html; https://publichealth.jhu.edu/2021/variants-vaccines-and-what-they-mean-for-covid-19-testing).
Table 3
|
Serology diagnostic assays and their sensitivity ranges, their target proteins, and several other key information. |
|||||
|
Type of test, source |
Kits available, Company, Approval |
Targets |
Sensitivity |
Quantitative or qualitative |
Pros and cons |
|
Total Ab-ECLIA (Datta et al., 2021; https://diagnostics.roche.com/us/en/products/params/elecsys-anti-sars-cov-2.html;https://diagnostics.roche.com/global/en/news-listing/2020/roche-develops-new-serology-test-to-detect-covid-19-antibodies.html) |
Elecsys AntiSARS-CoV-2 S, Roche Diagnostics, USFDA EUA |
SARS-CoV-2 Ag against spike (S) protein RBD |
≥ 14 days post PCR confirmation (95% Cl): 99.5% (97.0–100%) |
Quantitative |
Pros: Takes very little time (18 min), epidemiological application |
|
Lateral flow IgM/G (Datta et al., 2021; https://www.fda.gov/media/144071/download) |
Innovita 219-nCoV Ab Test (Colloidal Gold), Innovita (Tangshan) Biological Technology Co., Ltd., USFDA EUA |
SARS-CoV-2 Ag against S1 and nucleocapsid (N) proteins |
100% (30/30) with confidence interval of (88.7%, 100%). Possible lower sensitivity to IgG antibodies in symptomatic individuals less than 15 days since symptom onset. Sensitivity of this test after early infection is unknown |
Qualitative |
Pros: Aids with variant detection |
|
Total antibody, CLIA (Datta et al., 2021; https://www.fda.gov/media/138757/download) |
Dimension EXL SARS-CoV-2 Total antibody assay (CV2T), Siemens Healthcare Diagnostics Inc., USFDA EUA |
SARS-CoV-2 Ag against S1 protein RBD |
Sensitivity of this test after early infection is unknown |
Quantitative |
Pros: Low probability of false positives, detects total antibody levels |
|
IgM-CLIA (https://www.fda.gov/media/142911/download; https://www.beckmancoulter.com/products/immunoassay/access-sars-cov-2-igm-antibody-test; Datta et al., 2021) |
Access SARS-CoV-2 IgM, Beckman Coulter, Inc., USFDA EUA |
SARS-CoV-2 Ag against S1 protein RBD |
Sensitivity of this test after early infection is unknown |
Quantitative |
Pros: Low maintenance, easy workflow integration |
|
Total Ab-ELISA (Datta et al., 2021; https://www.thermofisher.com/covid-19-antibody-testing/us/en/solutions/OmniPATH-COVID19-Total-Antibody-ELISA-Test.html; https://www.fda.gov/media/142700/download) |
OmniPATH COVID-19 Total Antibody ELISA Test, Thermo Fisher Scientific, USFDA EUA |
SARS-CoV-2 Ag against S1 protein RBD |
≥ 15 days post symptom onset (95% Cl): 100% (89.2, 100.0). Sensitivity of this test after early infection is unknown |
Quantitative |
Pros: Detects IgM, IgA, IgG bound antibodies |
|
Total Ab-ELISA (Datta et al., 2021; https://www.fda.gov/media/137493/download; https://publichealth.jhu.edu/2021/variants-vaccines-and-what-they-mean-for-covid-19-testing) |
Platelia SARS-CoV-2 Total Ab assay, Bio-Rad, USFDA EUA |
SARS-CoV-2 Ag against the nucleocapsid (N) protein |
Overall: 49/50 (98%); 95% CI: 89.51–99.65% |
Quantitative |
Pros: One-step antigen capture (90 min incubation), visual control |
|
IgM-CLIA (Datta et al., 2021; https://www.diazyme.com/covid-19-antibody-tests/dz-lite-sars-cov-2-igm-clia-kit; https://www.fda.gov/media/141255/download) |
Diazyme DZ-Lite SARS-CoV-2 IgM CLIA Kit, Diazyme Laboratories, Inc., USFDA EUA |
SARS-CoV-2 Ag against the nucleocapsid (N) and spike (S) proteins |
Overall: 94.4% |
Quantitative |
Pros: High specificity, no cross-reactivity |
The Innovita 219-nCoV Ab Test (Colloidal Gold) employs a rapid lateral flow qualitative chromatographic immunoassay, and its sensitivity is about 100%, with its confidence interval of (88.7%, 100%) (https://www.centerforhealthsecurity.org/covid-19TestingToolkit/testing-basics/types-of-COVID-19-tests/serology-tests.html). This test uses antibodies against both the S1 and nucleocapsid (N) proteins of SARS-CoV-2 (https://www.fda.gov/media/144071/download), as the antigens from these regions are colloidal gold-labelled. Although mutations are possible in both regions, testing for antibodies that match two different proteins results in a higher efficacy.
The Dimension EXL SARS-CoV-2 Total antibody assay (CV2T) is an example of a quantitative chemiluminescent immunoassay, and its sensitivity after early SARS-CoV-2 infection in the host is unknown (https://www.centerforhealthsecurity.org/covid-19TestingToolkit/testing-basics/types-of-COVID-19-tests/serology-tests.html). The target antigen of this assay is the S1 protein, which has been associated with several possible mutations that could potentially lead to several COVID-19 variants (Fig. 1) (https://www.fda.gov/media/138757/download). Access SARS-CoV-2 IgM test also relies on a chemiluminescent immunoassay and its sensitivity after early infection is unknown and requires further study. This test can also yield false negative results if the quantity of antibodies against the SARS-CoV-2 virus is too low or if the virus has acquired one or more amino acid mutations in the RBD of the viral S1 protein recognized by the antibodies employed in this test. These challenges result in decreased assay efficacy (https://www.beckmancoulter.com/products/immunoassay/access-sars-cov-2-igm-antibody-test).
The OmniPATH COVID-19 Total Antibody Test is a quantitative enzyme-linked immunosorbent assay (ELISA), and its overall sensitivity is 100%, post 15 days since symptom onset (https://www.centerforhealthsecurity.org/covid-19TestingToolkit/testing-basics/types-of-COVID-19-tests/serology-tests.html). Similar to other serology tests, this test can also result in false negative results. Since this assay employs SARS-CoV-2 antibodies against the RBD of the S1 protein, its accuracy is also dependent on the possible mutations of this region in COVID-19 disease variants (https://www.thermofisher.com/covid-19-antibody-testing/us/en/solutions/OmniPATH-COVID19-Total-Antibody-ELISA-Test.html). The Platelia SARS-CoV-2 Total Ab assay is also an example of an enzyme-linked immunosorbent assay, and its overall sensitivity is 98%. This assay relies on SARS-CoV-2 antibodies against the nucleocapsid (N) protein (https://publichealth.jhu.edu/2021/variants-vaccines-and-what-they-mean-for-covid-19-testing).
Finally, the Diazyme DZ-Lite SARS-CoV-2 IgM CLIA Kit is a quantitative chemiluminescent immunoassay, like the Dimension EXL and Access SARS-CoV-2 IgM tests (https://www.centerforhealthsecurity.org/covid-19TestingToolkit/testing-basics/types-of-COVID-19-tests/serology-tests.html). Its overall sensitivity is approximately 94.4%. This assay uses antibodies targeted against nucleocapsid (N) and spike (S) proteins (https://www.diazyme.com/covid-19-antibody-tests/dz-lite-sars-cov-2-igm-clia-kit). By screening antibodies with two different target sites, as the Innovita 219-nCoV Ab Test does, the Diazyme DZ-Lite assay minimizes false negatives, and increases efficacy.
Within the larger realm of serology tests, there are two types: qualitative and quantitative antibody tests (https://www.centerforhealthsecurity.org/covid-19TestingToolkit/testing-basics/types-of-COVID-19-tests/serology-tests.html). Lateral flow assays (LFAs) are typically qualitative, rapid serology tests, which are used to provide a patient's “serostatus,” a common term denoting whether a patient has antibodies of interest. This test can be performed very rapidly at a single point in time, which is a major advantage. In contrast, quantitative tests include enzyme-linked immunosorbent assays (ELISAs) and chemiluminescent immunoassays (CLIAs), and similar tests provide additional detailed information beyond a binomial (detected or not detected) answer. Instead, these tests provide the levels of antibodies in a patient sample and are thus biologically quantitative, but are more heavily used for research purposes rather than informing patients.
Because these serology-based diagnostic tests rely on highly specific antibodies, serology assays can potentially result in false negatives in the presence of COVID-19 variants. Changes in the viral genome can result in downstream changes in the protein sequence and conformation. These changes can cause changes in the epitope region of the viral antigen, and these altered epitopes would specifically bind to antibodies with matching paratopes. The resulting antibody-antigen incompatibility due to mutation can result in false negatives and viral variants that spread while going undetected. Therefore, further development of serology-based diagnostic testing is needed to ensure accurate COVID-19 diagnoses in the face of COVID-19 variants.
3.3: CRISPR diagnostics
Clustered regularly interspaced short palindromic repeats (CRISPR)-Cas system is a genetic engineering technique that allows targeted modification of genomes. It is based on a simplified version of the bacterial CRISPR-Cas9 antiviral defence system (Palaz, Kalkan, Tozluyurt, & Ozsoz, 2021). Cas is an enzyme that can recognize and cleave specific target strands of DNA using a guide CRISPR sequence in prokaryotic organisms like bacteria and archaea (Ganbaatar & Liu, 2021). The CRISPR sequences are derived from DNA fragments of bacteriophages that had previously infected the prokaryote (Ganbaatar & Liu, 2021). The sequences allow the prokaryote to detect and destroy DNA from similar bacteriophages during subsequent infections, protecting the bacterial cell from invasion. Hence, these CRISPR sequences play a vital role in the antiviral defence of prokaryotes and provide a form of acquired immunity.
In general, there are two major parts in the CRISPR-Cas system: guide RNA (to identify and direct Cas endonuclease to the target region) and Cas endonuclease (to break the target genomic site) (Ding et al., 2020). By delivering the Cas9 nuclease complexed with a synthetic guide RNA into a cell, the genome can be cleaved at the desired location, allowing existing genes to be removed and new ones introduced in vitro or in vivo (in living organisms) (Ganbaatar & Liu, 2021).
3.3.1: CRISPR and COVID-19 detection
CRISPR-based detection methods have recently received substantial attention for nucleic acid-based molecular testing due to their simplicity, high sensitivity, and specificity. There are various CRISPR-based COVID-19 detection methods and related diagnostic devices. As of August 2021, over 217 million people worldwide have been infected, with over four million deaths due to this virus. Thus, there is a critical need for simple, rapid, and affordable testing facilities in every country, from developing to first-world nations. With the help of CRISPR, proteins can precisely cut the target region that matches the complementary crRNA sequence (Ganbaatar & Liu, 2021). There are two components in CRISPR detection: first, the CRISPR-RNA complex cuts the target region. This step initiates the next one, collateral cleavage of the surrounding nucleic acids. Generally, the CRISPR system is divided into two main classes and six types (Broughton et al., 2020a). Class I contains Cas proteins that cut DNA and RNA in vivo. Class II CRISPR systems are used widely for genomic manipulation and infectious disease diagnosis, and these systems feature three main effector proteins: Cas12, Cas13, and Cas14 (Broughton et al., 2020a).
Concerning cleavage activity, Cas12 recognizes double-stranded DNA (dsDNA) more efficiently than single-stranded DNA (ssDNA), but it still exhibits collateral activity for ssDNA (Broughton et al., 2020a). Cas14 recognizes ssDNA more effectively than dsDNA as well, and Cas14 also exhibits similar collateral activity in ssDNA. Cas13 possesses the intriguing ability to both recognize ssRNA and exhibit collateral activity. Since CRISPR can create collateral cleavage activity, it can be combined with isothermal nucleic acid amplification to simplify the detection method by visualizing the result of positive or negative samples with the naked eye, LED or UV lamps (Broughton et al., 2020a).
There is another test to diagnose COVID-19; however, it is not as popular (Rahimi et al., 2021). A multiplex diagnostic system developed recently incorporated nucleic acid preamplification with CRISPR/Cas enzymology to identify the targeted nucleic acid sequences with high sensitivity (Kellner, Koob, Gootenberg, Abudayyeh, & Zhang, 2019). The developed system, called specific high-sensitivity enzymatic reporter unlocking (SHERLOCK) can detect clinical sample nucleic acid sequences in a portable and ultrasensitive manner.
In recent months, Cas12 has been used in several assays. One commonly used CRISPR test is the DETECTR (SARS-CoV-2 DNA Endonuclease-Targeted CRISPR Trans Reporter) (Kaminski, Abudayyeh, Gootenberg, Zhang, & Collins, 2021). This assay uses loop-mediated amplification (RT-LAMP) to reverse-transcribe and amplify viral RNA obtained from a nasal or oral swab. The Cas12 protein then identifies target SARS-CoV-2 sequences, leading to genetic cleavage that signals a positive test result (Rahimi et al., 2021). The entire test takes only 30–40 min to complete, and the results can be seen using a lateral flow strip. If both the E and N protein-encoding genes are detected, the DETECTR assay signals a positive result. However, the result changes to presumptive positive if only one of the two protein-encoding genes is detected (Rahimi et al., 2021).
The FAM-biotin reporter molecule and lateral flow strips provide a visual result for the Cas12 detection. Flow strips capture and separate the cleaved strands from the uncleaved reporter molecules (Rahimi et al., 2021). The RT–LAMP DETECTR assay's ability to amplify SARS-CoV-2 nucleic acid directly from the swab sample is a valuable and critical component of the diagnostic's efficacy and usability. Broughton et al. performed a study where they discovered diminishing accuracy from the diagnostic with higher reaction concentrations of ≥ 10% UTM and ≥ 10% phosphate-buffered saline by volume, with estimated LODs decreasing to 15,000 and 500 copies per μL, respectively (Broughton et al., 2020a; Rahimi et al., 2021).
CRISPR-based diagnostic systems have reshaped molecular diagnosis. The benefits of the CRISPR system, such as speed, precision, specificity, strength, efficiency, and versatility, have inspired researchers to develop CRISPR-based diagnostic and therapeutic methods (Rahimi et al., 2021). For instance, eliminating the need for thermocycling and isothermal signal amplification offers significant benefits compared to qRT-PCR, such as fast turnaround time, target specificity for single nucleotides, integration with usable and user-friendly reporting formats like lateral flow strips, and no requirement for sophisticated laboratory systems (Rahimi et al., 2021). During the global COVID-19 outbreak, different groups have begun designing and developing diagnostic and therapeutic programs based on the efficient CRISPR system. Thanks to these developments, scientists will be able to accurately run cost-effective tests and apply these methods to other groundbreaking molecular work.
3.4: Biosensor diagnostics
A biosensor is a device that is assembled by utilizing two major components: the biorecognition element and the transducer within. The former enables the detection of an analyte, and the latter translates this specific interaction into a quantifiable signal. The whole process of recognition is referred to as “signalization.” Transducers have been classified according to the type of signal generated upon analyte-bioreceptor interaction. Biosensors are considered one of the most sensitive, specific and efficient diagnostics for disease detection, and consequently, they have been harnessed to monitor COVID-19 spread. Sensors for SARS-CoV-2 were designed based on viral surface nucleoproteins, receptors, and genetic material. Biosensors can be broadly classified into four categories: (1) optical, (2) thermal, (3) electrochemical, and (4) piezoelectric. In the following sections, we discuss the types of biosensors that are prevalent and their conceptual operation in SARS-CoV-2 diagnostics.
3.4.1: Optical biosensors
Optical biosensors mainly work by converting light signals into electrical signals. The activity is based on the interaction of the optical field with a bio-recognition element. These sensors have been categorized as “label-free” and “label-based.” The first approach generates signals directly upon the interaction of the analyte material with the transducer. On the other hand, signals produced in label-based sensing are amplified by colorimetric, fluorescent or luminescent methods. Optical sensors have specialized into different categories such as surface plasmon resonance (SPR), evanescent wave fluorescence, and optical waveguide interferometry (Table 4).
Table 4
|
COVID-19 biosensors designed for rapid detection of disease. |
|||
|
Type of sensor |
Detection |
LOD or sensitivity |
Reference |
|
FET |
Sensor detects SARS-CoV2 antigen protein. Graphene base functionalized with SARS-CoV2 spike antibody used |
1 fg/mL |
Seo et al. (2020) |
|
Electrochemical |
SARS-CoV-2 antigen is detected with the potentiostat by measuring the change in electrical conductivity. The SARS-CoV2 monoclonal antibody was immobilized onto a screen printed carbon electrode (SPCE) |
90 fM within 10–30 s |
Mahari, Roberts, Shahdeo, and Gandhi (2020) |
|
Electrochemical |
N and S genes of SARS-CoV2 detected in less than 2 h. RCA is used to generate amplicons which are hybridized with probes functionalized with redox active labels |
1 copy/ μL |
Chaibun et al. (2021) |
|
Electrical |
SARS-CoV2 S1 spike protein detected with the help of Bioelectric Recognition Assay. The antigen specific antibody is bound to membrane-engineered mammalian cells. As the protein attaches to the antibody considerable change is observed in the cellular bioelectric properties |
1 fg/mL |
Mavrikou, Moschopoulou, Tsekouras, and Kintzios (2020) |
|
Electrochemical |
SARS-CoV2 antibodies are detected by the sensor. Here, the presence of Ab stops a redox conversion which leads to a decrease in current that is measured |
IgG = 0.96 ng/mL |
Yakoh et al. (2021) |
|
Electrochemical |
RNA of SARS-CoV-2 is detected using calixarene functionalized graphene oxide which targets the RNA without any requirement of nucleic acid amplification and reverse-transcription. Only portable electrochemical smartphones are a necessity |
200 copies/mL |
Zhao et al. (2021) |
|
LFIA (optical) |
SARS-CoV2 nucleocapsid protein was detected using this platform. Specific single chain variable fragment- crystallizable fragment (scfc-fc) fusion antibodies were developed using phage display technique. Cellulose nano beads were used for visual display of attached SARS antigen |
2 ng of antigen protein; |
Kim et al. (2021) |
|
Raman scattering based LFIA |
Simultaneous detection of anti SARS-CoV2 IgM/IgG is performed. Dual-layer Raman molecules loaded silver-coated SiO2 nanoparticles as SERS tags in clinical samples. The Raman molecule used was DTNB; system surface was modified with CoV2 spike protein which specifically binds antibodies |
1 pg/mL |
Liu et al. (2021) |
|
SPR |
Dual functional plasmonic biosensor which combines the plasmonicphotothermal effect and localized surface plasmon resonance detects the CoV2 specific genes. 2D Au-Nanoislands were functionalized with complementary DNA receptors which specifically bind to Cov2 sequences and generate thermoplasmonic heat |
0.22 pM |
Qiu et al. (2020) |
|
SPR |
SPR sensor developed for the detection of nucleocapsid protein antibodies from serum samples. The sensor is coated with the recombinant nucleocapsid protein which specifically binds to the antibody from the serum. |
nM range within 15 min |
Djaileb et al. (2020) |
|
Optical |
Split luciferase mechanism used for antibody detection specific to S protein and N protein of SARS-CoV2. NanoLuc was split into two and both arms fused with viral antigens. The antibodies bind with both the arms and reconstruct the NanoLuc, hence generating the luminescence |
89% for S protein; |
Elledge et al. (2021) |
|
Colorimetric and SPR |
Detection of N gene of SARS-CoV2 by the use of gold nanoparticles capped with thiol-modified antisense oligonucleotides that are N gene specific. The binding of the target demonstrates a change in surface plasmon resonance. The cleavage of RNA-DNA hybrid gives a detectable precipitate because of addition of RNase H |
0.18 ng/mL |
Moitra, Alafeef, Dighe, Frieman, and Pan (2020) |
|
Optical (LFIA, Fluorescence) |
Single stranded Recombinase Polymerase Amplification method used for detection of SARS-CoV2 RNA. Isothermal amplification of RPA is done and then dsDNA converted to a single strand which can be detected using the hybridization process within 10 min |
4 copies/50 μL sample |
Kim et al. (2020) |
|
Optomagnetic |
Conserved region of SARS-CoV2 RNA dependent RNA polymerase (RdRp) gene detected by making synthetic cDNA and applying the circle to circle amplification; a rolling circle amplification based cascade |
0.4 fM |
Tian, Gao, Fock, Dufva, and Hansen (2020) |
|
Fluorescence and LFA |
E gene and N gene of the SARS-CoV2 are targeted in this assay. Simultaneous reverse transcription and isothermal amplification is done using LAMP. Later CRISPR technique is used for the detection wherein Cas12 g RNA are utilized targeting the N and E gene |
10 copies/μL |
Broughton et al. (2020b) |
|
Fluorescence and LFA |
CASdetec is a developed nucleic acid detection platform based on CRISPR technique for the detection of RdRp of SARS-CoV2 using sgRNA-3. Recombinase aided amplification is used to amplify the substrates |
1 × 104 copies/mL |
Guo et al. (2020) |
|
Fluorescence and LFA |
Multiplex reverse transcription loop-mediated isothermal amplification (mRT-LAMP) is designed for the detection of SARS-CoV2 combined with nanoparticle-based lateral flow biosensor. The opening reading frame 1a/b and N gene of the virus were detected |
12 copies per reaction with 100% sensitivity |
Zhu et al. (2020) |
|
Fluorescence, colorimetric and LFA |
Immunoassay with fluorescence-colorimetry dual mode LFIA for detection of IgM and IgG specific to SARS-CoV2 from serum. Spike protein is used for the detection which is conjugated with SiO2@Au@QD nanobeads |
1:106 dilution by fluorescence values |
Wang et al. (2020) |
|
Optical and LFIA |
Biolayer interferometry technique is used for the detection of spike protein wherein α,N-acetyl neuraminic acid was immobilized onto sensor platform and signal was generated upon binding of spike protein to glycan |
5 μg/mL |
Baker et al. (2020) |
|
Optical and LFIA |
SARS-CoV2 IgG and IgM were detected from serum, plasma and whole blood. Colloidal-gold labelled SARS-CoV-2 antigen was used for detection |
92% sensitivity for venous blood; |
Black et al. (2021) |
|
Fluorescence and LFIA |
Detection of anti SARS-CoV2 IgG antibody in human serum. The capture molecule was the recombinant SARS-CoV2 nucleocapsid phosphoprotein. The detection was done using Lanthanide-doped polystyrene nanoparticles which served as a fluorescence reporter functionalized with IgG mouse/rabbit anti-human IgG antibody |
– |
Chen et al. (2020) |
|
LFIA |
The method developed for simultaneous detection of IgM and IgG specific to SARS-CoV2. The surface antigen conjugated with gold nanoparticles is used here as a capture |
Sensitivity = 88.63% |
Li et al. (2020) |
|
Optical (Interferometry) |
The sensor is developed based on an interferometric bimodal waveguide whose surface is modified with specific receptors targeting antigens of SARS-CoV2 such a spike protein. When virus particles are captured by receptors, interferometric signals are recorded |
Sensitivity = greater than 95% |
Ruiz-Vega, Soler, and Lechuga (2021) |
|
Plasmonic Sensor |
Optical detection of SARS-CoV2 virus done using the specific monoclonal antibody and ACE2 protein |
370 virus particles/mL |
Huang et al. (2021) |
|
LFA |
Evaluation of four lateral flow assay kits for detection of SARS-CoV2 IgG |
Sensitivities: |
McAulay et al. (2020) |
|
LFIA |
Detection method for SARS-CoV2 spike 1 protein using a combination of capture and detector molecule being SARS-CoV2 receptor ACE2 and commercially available antibodies for S1 respectively |
1.86 × 105 copies/mL |
Lee et al. (2021) |
|
Quartz Crystal Microbalance (Piezo) |
S1 spike protein of SARS-CoV2 is detected with the help of anti-spike glycoprotein (engineered surface with mixed SAM of CH3 and COOH + antibody) |
40 nM |
Pandey (2020) |
|
Amplification and Microfluidic device |
SARS-CoV2 N gene and SARS-CoV2 S gene are being detected using isothermal amplification analyser (RTisochip). The chip is able to detect 19 common respiratory viruses as well |
S gene = 10 copies/μL; |
Xing et al. (2020) |
|
Chromatographic immunoassay |
Standard Q COVID-19 antigen (SD Biosensor) for detection of SARS-CoV2 nucleoprotein in nasopharyngeal swabs |
Sensitivity = 70.6% |
Cerutti et al. (2020) |
SPR is a type of label-free optical detection whereby the transducer consists of a probe attached to a thin metallic film, which remains in close contact with a dielectric medium of lower refractive index. Subsequent binding of the analyte changes the angle of extinction of light, which is reflected after transverse waves impinge on the metal dielectric interface. This fluctuation may be perceived as a signal and is directly proportional to the concentration of analyte present on the surface (Shrivastav, Cvelbar, & Abdulhalim, 2021; Unser, Bruzas, He, & Sagle, 2015). Djaileb et al. developed a SPR based sensor for the detection of antibodies against the N protein from serum (Djaileb et al., 2020). Similarly, Qiu et al. achieved detection of various genes specific to COVID-19 with the introduction of dual functional plasmonic sensors (Qiu et al., 2020). Table 4 exhaustively lists the methods that have applied this technology.
3.4.2: Electrochemical biosensors
Electrochemical sensors use electrochemical transducers to detect the inherent electrical properties of interacting biomolecules. Electrochemical mechanisms generally require sensing or redox electrodes which include a reference electrode, a counter electrode, and a working electrode in combination. The working electrode provides the interface where the probe is immobilized. It further acts as a transduction element for the elicitation of electrical fluctuations. The counter electrode functions to maintain a continuum with the electrolyte solution, thus maintaining the circuit.
Electrochemical sensors have been mainly classified into three types: amperometric, potentiometric, and conductometric devices. Amperometric devices measure the current, caused by oxido-reduction reactions of an electroactive molecule, at a constant potential. In contrast, voltammetry is the term used for the current measured with controlled variation in the potential. Not all analytes are intrinsically capable of inducing redox reactions; in such cases external mediators, also known as indicators, facilitate the electrochemistry of the analyte at the working electrode surface. Potentiometric devices mainly indicate the concentration of ions related to electrochemical reactions by measuring the charge potential that is accumulated on the working electrode. Hence, the electromotive force defines the relationship between the concentration and the potential at the working electrode, which is also given by the Nernst equation. Conductometric devices work by measuring the ability of an analyte to conduct an electrical current between electrodes. They are contemplated as a subset of impedimetric devices. These devices are widely used for electrochemical reactions using enzymes, wherein the ionic strength of the conductive solution may change due to the reaction in situ.
Huge preference for electrochemical transducers is attributed to ease of synthesis, quick response time, and low power consumption (Table 4) (Desai, Kumar, Bose, & Datta, 2018; Grieshaber, MacKenzie, Vörös, & Reimhult, 2008). Seo and his associates have developed an electrochemical sensor capable of detecting the S protein with a sensitivity of 1 fg/mL, whereas Mahari et al. developed a similar sensor with a sensitivity up to 90 fM (Seo et al., 2020). Later in 2021, Zhao et al. fabricated a technique for the electrochemical detection of SARS-CoV2 RNA that did not require nucleic acid amplification (Zhao et al., 2021). Variants and modified electrochemical sensors have been actively used for COVID19 detection (Table 4).
3.4.3: Piezoelectric biosensors
Piezoelectric biosensors are mainly designed for the detection of affinity interactions. These biosensors elicit a voltage when they are induced due to mechanical or oscillatory stress. Alternatively, voltage applied to piezoelectric material causes oscillation or mechanical stress on the crystal, whose frequency may be detected by putting the crystal into an oscillation circuit. The detection module of the sensors are located on the surface of the crystal, and binding of the analyte causes fluctuations in the oscillation frequency. This frequency observed is directly proportional to the analyte concentration (Hussain, Rupp, Wendel, & Gehring, 2018). Among the many sensors created, the quartz crystal microbalance developed by Pandey specifically detects the S1 spike protein of SARS-CoV2 at protein levels as low as 40 nM (Pandey, 2020).
3.4.4: Next generation sensors
Wearable sensors are considered as a class of next gen sensor, having application in both diagnostics and regular monitoring. The magnitude of wearable sensors is vast and varies from physiological sensing and biochemical sensing to motion sensing. The ability to miniaturize the electronic circuits and hence miniaturize the sensor plays a major role in the development of wearable sensors. The applicability of accelerometers has been shown to help monitor the activity of daily living, especially for old people. Wearable sensors can help track for years the recovery of patients who have undergone abdominal surgery. These types of sensors are now preferred even more due to the advantages they offer, especially since no sampling or processing is required. Thus, these next gen sensors can be easily used by the general population.
With face masks becoming a necessity and a preventive safety measure, scientists have delved into the plausibility of sensor-based face masks as a strategy for identifying infected individuals. Researchers from Harvard and MIT designed a SARS-CoV-2 sensing mask capable of detecting a patient within 90 min. Embedded with miniature sensors, the technology encompasses freeze-dried cellular machinery capable of detecting viral particles in the exhaled aerosol of the person wearing the mask. This sensor-based mask has a sensitivity equivalent to that of gold-standard WHO-approved RT-PCR tests, and it can detect the presence of the viral genome. This particular novel prototype has begun another path for the next generation sensors. As of now, the prototype biosensor costs approximately $5, and it is anticipated to become cheaper with mass manufacturing (Nguyen et al., 2021).
Another upcoming and explorative field in next generation sensors are CRISPR-based sensors. The ability to target several genomic loci simultaneously is achieved with the help of CRISPR and its associated proteins known as Cas proteins, as mentioned previously. With CRISPR being one of the most promising potential fields of research, the detection of many pathogens are being targeted with the help of CRISPR, including the detection of coronaviridae. As described before, CRISPR-based SHERLOCK assays are being utilized specifically for the detection of SARS-CoV-2. This detection can be achieved by using samples from nasal swabs, nasopharyngeal swabs, oropharyngeal swabs or bronchoalveolar lavage (BAL). As the viral signature sequence is detected, the CRISPR enzyme is released to generate fluorescent signals (Azhar et al., 2020). A CRISPR-based in vitro diagnostic (IVD) detector is used for the detection of E and N2 genes of SARS-CoV-2 by using Cas12a. The Cas12a-sgRNA complex binds to the target, leading to ssDNA cleavage and concurrent increase in fluorescence (Kellner et al., 2019). CRISPR-based detection offers the user the sensitivity and specificity they require in detection of viruses belonging to the same family, and it is robust enough to distinguish between genetically similar viruses. This advantage is attributed to the fact that detection is based on specific signature sequences that are conserved (Datta et al., 2021).
4: Diagnostics in the era of COVID-19 vaccination
While vaccines are indispensable in the fight against COVID-19, it is essential to consider how vaccination alters viral diagnostics, specifically whether SARS-CoV-2 vaccination results in a false-positive diagnosis. There are two broad categories of testing: viral tests and serology (antibody) tests. There are two broad categories of testing: viral tests and serology (antibody) tests. Viral tests include antigen tests (often referred to as rapid tests) and nucleic acid amplification tests (NAAT). Briefly, NAATs (e.g. RT-PCR) detect viral RNA, whereas rapid antigen tests detect specific viral proteins (i.e. antigens) on the virus' surface. In contrast, serology tests, described interchangeably as antibody tests, measure specific viral antibody levels. These serology tests can also be performed rapidly, though they only provide information on antibody presence, not viral infection.
Both viral tests are highly unlikely to result in a false-positive test because of vaccination for two primary reasons. First, mRNA COVID-19 vaccines (mRNA-1273 by Moderna and BNT162b2/Comirnaty by Pfizer-BioNTech) and adenovirus-based COVID-19 vaccines (ChAdOx1-S by AstraZeneca-Oxford and Janssen COVID-19 Vaccine by Johnson & Johnson) do not consist of the entire SARS-CoV-2 genome. Instead, these four vaccines contain a virus fragment that carries instructions on synthesizing S-proteins. Therefore, these vaccines use the S-protein as a major target. Fortunately, the majority of widely used molecular tests do not target the S gene. Moreover, S gene-targeting molecular tests (e.g. Thermofisher Taqpath) have other gene targets, thereby remaining functionally reliable. Thus, vaccination does not significantly affect the accuracy of viral tests (Borges et al., 2021).
Second, mRNA-1273 (Moderna) and BNT162b2/Comirnaty (Pfizer-BioNTech) contain non-replicating mRNA. Specifically, these two vaccines contain the target antigen sequence, 3′ untranslated region (UTR), and 5′ UTR but lack protein sequences necessary for self-amplification. Consequently, these non-replicating viral vector vaccines are unable to create new viral particles upon host entry. In other words, the small amount of mRNA in these non-replicating mRNA vaccines is too low to be detected by either RT-PCR or antigen testing (Guglielmi, 2020).
On the other hand, vaccination can result in false positive results for serology (antibody) tests. A false positive can occur if the serology test specifically detects spike protein-specific antibodies. All four vaccines mentioned in the previous paragraph (i.e. mRNA-1273, BNT162b2/Comirnaty, ChAdOx1-S, and Janssen COVID-19 vaccine) aim to elicit antibody production against the spike protein. Hence, a positive result from serology tests that target the spike protein (Table 3) could indicate either vaccination or prior infection. In contrast, serology tests that target antibodies to the nucleocapsid protein (Table 3) provide more conclusive results; a positive test result could only occur from natural infection, allowing for differentiation between natural infection and immunization (West, Gronvall, & Kobokovich, 2021).
Another question is whether serology tests are appropriate diagnostic tools to measure vaccine-induced immunity against SARS-CoV-2. Unfortunately, there is a lack of research that evaluates the level of protection provided by an immune response to COVID-19 vaccination. As of October 2021, the FDA has not recommended any currently available serology tests as a reliable method to measure vaccination efficacy.
Serology testing remains an unreliable predictor of COVID-19 immunity for two main reasons. First, scientists and clinicians have yet to determine the exact concentration of antibodies needed to prevent SARS-CoV-2 infection or illness (correlates of protection). Due to this gap of knowledge, a positive serology test obtained via qualitative or quantitative COVID-19 serology testing, including those listed in Table 3, does not guarantee protection against COVID-19. Second, as listed in Table 3, numerous serology tests target antibodies to the N protein rather than the S protein. Since currently approved COVID-19 vaccines elicit S protein-specific neutralizing antibodies, people who have had the COVID-19 vaccine (but not a natural infection) would test negative with serology tests that do not detect anti-S protein antibodies. Therefore, serology tests are not an accurate measure of COVID-19 vaccine efficacy. Importantly, misinterpretation of serology tests can provide a false sense of security, misleading people away from clinically proven security measures such as mask wearing and social distancing (Bausch, Hampton, Perkins, & Saville, 2021).
5: Conclusion
At the time of writing, the COVID-19 pandemic continues, causing widespread infection and death. Diagnostic techniques involving RT-PCR, serology, CRISPR, and biosensors have all played roles in informing best health practices and slowing the spread of SARS-CoV-2. However, the disease has become more transmissible, dangerous, and challenging to detect with the increasing number of emerging variants. Developments in diagnostic methods have helped, but the spread of the virus continues to rage. People continue to lose their lives daily, despite the world's best diagnostic capabilities. Many countries cannot afford the best diagnostic assays, resulting in health inequities on a global scale.
It must be mentioned that diagnostics alone are incapable of bringing an end to the COVID-19 pandemic. While testing helps mitigate the spread, the best way to stop the spread of COVID-19 is global vaccination, augmenting acquired immunity against the virus and limiting viral transmission (and severe infection). Vaccination is crucial to curb variant emergence and transmission frequency while preventing further viral replication and mutation (Salian et al., 2021). Effective vaccine distribution reduces the need for diagnostic testing since fewer individuals will transmit and develop viral infections, reducing the population's viral load. This reduction in testing demand is vital, given the immense strain that diagnostic efforts have put on the global scientific and clinical supply chains. Therefore, vaccines are necessary to stop the spread of COVID-19, mitigate the need for testing, and save lives. Coupled with implemented public safety measures such as physical distancing, vaccinations provide hope for a safer and healthier post-pandemic future (Ramos, Vela-Pérez, Ferrández, Kubik, & Ivorra, 2021).
Unfortunately, just as the distribution of testing was not always equal, neither has the worldwide distribution of vaccines. Many countries have been left out of the benefits of vaccination because they lacked resources and global support (Salian et al., 2021). Thus, people in these countries must face the dangerous COVID-19 variants without protection from vaccines. This predictable and sad reality is worsened by vaccine hesitancy and mistrust within countries with greater vaccine access (Forni & Mantovani, 2021). These factors contribute to the massive health disparities related to vaccination worldwide. According to the New York Times, over 5 billion vaccine doses have been administered worldwide. Yet, billions of people remain unvaccinated while health disparities worsen. Countries like the United States, France, China, and Canada have over 100 vaccine doses administered per 100 people. In contrast, countries like Venezuela, Nicaragua, Kenya, and South Africa average fewer than 20 doses per 100 people (Holder, 2021). These statistics point to a worldwide issue of access to healthcare that has existed throughout the COVID-19 pandemic.
With continued massive testing efforts, valuable resources remain in short supply, slowing down research and science while infections and deaths persist. The battle against COVID-19 continues, and a global solution is needed to end it. A worldwide collaborative effort to continue developing diagnostic assays is crucial to keeping up with the ever-changing virus and its variants. Researchers and scientists must cooperate and refine diagnostics that are accurate and sensitive to emerging SARS-CoV-2 variants so new variants can be identified and contained quickly. Further, these improved diagnostics should be made available to all people: ensuring the distribution of diagnostic tests to developing countries is a global responsibility. A globally run task force devoted to monitoring SARS-CoV-2 spread worldwide in real-time would provide a much-needed solution to the underreported COVID-19 cases around the world. To reiterate an earlier point, equitable vaccine distribution to people in every nation is also a global responsibility. With a worldwide cooperative effort, future waves of cases caused by new variants can be quickly contained and ultimately prevented with the proper investment in global accessibility for COVID-19 testing and vaccination.
References
Abdool Karim and de Oliveira, 2021 Abdool Karim S.S., de Oliveira T. New SARS-CoV-2 Variants—Clinical, public health, and vaccine implications. The New England Journal of Medicine. 2021;384(19):1866–1868. doi:10.1056/NEJMc2100362.
Arevalo-Rodriguez et al., 2020 Arevalo-Rodriguez I., Buitrago-Garcia D., Simancas-Racines D., Zambrano-Achig P., Campo R.D., Ciapponi A., et al. False-negative results of initial RT-PCR assays for COVID-19: A systematic review. PLoS One. 2020;15(12):e0242958doi:10.1371/journal.pone.0242958.
Azhar et al., 2020 Azhar M., Phutela R., Ansari A.H., Sinha D., Sharma N., Kumar M., et al. Rapid, field-deployable nucleobase detection and identification using FnCas9. 2020.doi:10.1101/2020.04.07.028167.
Babb de Villiers, Blackburn, Cook, & Janus, 2021 Babb de Villiers Chantal, Blackburn Laura, Cook Sarah, Janus JoannaPHG Foundation. SARS-CoV-2 variants. Foundation for Innovative New Diagnostics. 2021. https://www.finddx.org/wp-content/uploads/2021/03/COVID-variants-report-FINAL-12MAR2021.pdf.
Baker et al., 2020 Baker A.N., Richards S.-J., Guy C.S., Congdon T.R., Hasan M., Zwetsloot A.J., et al. The SARS-COV-2 spike protein binds sialic acids and enables rapid detection in a lateral flow point of care diagnostic device. ACS Central Science. 2020;6(11):2046–2052. doi:10.1021/acscentsci.0c00855.
Bausch, Hampton, Perkins and Saville, 2021 Bausch, D., Hampton, L., Perkins, M., & Saville, M. (2021). COVID-19: Why we can't use antibody tests to show that vaccines are working. https://www.gavi.org/vaccineswork/covid-19-why-we-cant-use-antibody-tests-show-vaccines-are-working
BioRender.com, 2021 BioRender.com. https://biorender.com.
Black et al., 2021 Black M.A., Shen G., Feng X., Garcia Beltran W.F., Feng Y., Vasudevaraja V., et al. Analytical performance of lateral flow immunoassay for SARS-CoV-2 exposure screening on venous and capillary blood samples. Journal of Immunological Methods. 2021;489(112909):doi:10.1016/j.jim.2020.112909.
Borges et al., 2021 Borges V., Sousa C., Menezes L., Gonçalves A.M., Picão M., Almeida J.P., et al. Tracking SARS-CoV-2 lineage B.1.1.7 dissemination: Insights from nationwide spike gene target failure (SGTF) and spike gene late detection (SGTL) data, Portugal, week 49 2020 to week 3 2021. Eurosurveillance. 2021;26 (10):doi:10.2807/1560-7917.ES.2021.26.10.2100130.
Broughton et al., 2020a Broughton J.P., Deng X., Yu G., Fasching C.L., Servellita V., Singh J., et al. CRISPR-Cas12-based detection of SARS-CoV-2. Nature Biotechnology. 2020a;38:870–874. doi:10.1038/s41587-020-0513-4.
Broughton et al., 2020b Broughton J.P., Deng X., Yu G., Fasching C.L., Singh J., Streithorst J., et al. Rapid detection of 2019 novel coronavirus SARS-CoV-2 using a CRISPR-based DETECTR lateral flow assay. medRxiv : the preprint server for health sciences. 2020b.doi:10.1101/2020.03.06.20032334 Submitted for publication.
Bustin, Benes, Nolan and Pfaffl, 2005 Bustin S.A., Benes V., Nolan T., Pfaffl M.W. Quantitative real-time RT-PCR – A perspective. Journal of Molecular Endocrinology. 2005;34(3):597–601. doi:10.1677/jme.1.01755.
CDC, 2021 CDC. CDC diagnostic tests for COVID-19. Centers for Disease Control and Prevention; 2021, August 7. https://www.cdc.gov/coronavirus/2019-ncov/lab/testing.html.
Cerutti et al., 2020 Cerutti F., Burdino E., Milia M.G., Allice T., Gregori G., Bruzzone B., et al. Urgent need of rapid tests for SARS CoV-2 antigen detection: Evaluation of the SD-biosensor antigen test for SARS-CoV-2. Journal of Clinical Virology: The Official Publication of the Pan American Society for Clinical Virology. 2020;132:104654doi:10.1016/j.jcv.2020.104654.
Chaibun et al., 2021 Chaibun T., Puenpa J., Ngamdee T., Boonapatcharoen N., Athamanolap P., O'Mullane A.P., et al. Rapid electrochemical detection of coronavirus SARS-CoV-2. Nature Communications. 2021;12(1):802. doi:10.1038/s41467-021-21121-7.
Chen et al., 2020 Chen Z., Zhang Z., Zhai X., Li Y., Lin L., Zhao H., et al. Rapid and sensitive detection of anti-SARS-CoV-2 IgG, using lanthanide-doped nanoparticles-based lateral flow immunoassay. Analytical Chemistry. 2020;92(10):7226–7231. doi:10.1021/acs.analchem.0c00784.
Cheng et al., 2021 Cheng L., Song S., Zhou B., Ge X., Yu J., Zhang M., et al. Impact of the N501Y substitution of SARS-CoV-2 spike on neutralizing monoclonal antibodies targeting diverse epitopes. Virology Journal. 2021;18(1):87. doi:10.1186/s12985-021-01554-8.
Cherian et al., 2021 Cherian S., Potdar V., Jadhav S., Yadav P., Gupta N., Das M., et al. SARS-CoV-2 spike mutations, L452R, T478K, E484Q and P681R, in the second wave of COVID-19 in Maharashtra, India. Microorganisms. 2021;9(7):1542. doi:10.3390/microorganisms9071542.
Datta, Singh and Naqvi, 2021 Datta M., Singh D.D., Naqvi A.R. Molecular diagnostic tools for the detection of SARS-CoV-2. International Reviews of Immunology. 2021;40(1–2):143–156. doi:10.1080/08830185.2020.1871477.
Dejnirattisai et al., 2021 Dejnirattisai W., Zhou D., Supasa P., Liu C., Mentzer A.J., Ginn H.M., et al. Antibody evasion by the P.1 strain of SARS-CoV-2. Cell. 2021;184(11):doi:10.1016/j.cell.2021.03.055 2939–2954.e9.
Desai, Kumar, Bose and Datta, 2018 Desai D., Kumar A., Bose D., Datta M. Ultrasensitive sensor for detection of early stage chronic kidney disease in human. Biosensors & Bioelectronics. 2018;105:90–94. doi:10.1016/j.bios.2018.01.031.
Deyde, Sampath and Gubareva, 2011 Deyde V.M., Sampath R., Gubareva L.V. RT-PCR/electrospray ionization mass spectrometry approach in detection and characterization of influenza viruses. Expert Review of Molecular Diagnostics. 2011;11(1):41–52. doi:10.1586/erm.10.107.
Di Giacomo, Mercatelli, Rakhimov and Giorgi, 2021 Di Giacomo S., Mercatelli D., Rakhimov A., Giorgi F.M. Preliminary report on severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike mutation T478K. Journal of Medical Virology. 2021;93(9):5638–5643. doi:10.1002/jmv.27062.
Ding et al., 2020 Ding X., Yin K., Li Z., Lalla R.V., Ballesteros E., Sfeir M.M., et al. Ultrasensitive and visual detection of SARS-CoV-2 using all-in-one dual CRISPR-Cas12a assay. Nature Communications. 2020;11(1):4711. doi:10.1038/s41467-020-18575-6.
Djaileb et al., 2020 Djaileb A., Charron B., Jodaylami M.H., Thibault V., Coutu J., Stevenson K., et al. A rapid and quantitative serum test for SARS-CoV-2 antibodies with portable surface plasmon resonance sensing. ChemRxiv; 2020 Submitted for publication.
Dyer, 2021 Dyer O. Covid-19: Indonesia becomes Asia's new pandemic epicentre as delta variant spreads. BMJ. 2021;n1815:doi:10.1136/bmj.n1815.
Elledge et al., 2021 Elledge S.K., Zhou X.X., Byrnes J.R., Martinko A.J., Lui I., Pance K., et al. Engineering luminescent biosensors for point-of-care SARS-CoV-2 antibody detection. Nature Biotechnology. 2021;39(8):928–935. doi:10.1038/s41587-021-00878-8.
Farasani, 2021 Farasani A. Biochemical role of serum ferratin and d-dimer parameters in COVID 19 diagnosis. Saudi Journal of Biological Sciences. 2021;doi:10.1016/j.sjbs.2021.08.040.
Faria et al., 2021 Faria N.R., Mellan T.A., Whittaker C., Claro I.M., da Candido D.S., Mishra S., et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science. 2021;372(6544):815–821. doi:10.1126/science.abh2644.
Fomsgaard and Rosenstierne, 2020 Fomsgaard A.S., Rosenstierne M.W. An alternative workflow for molecular detection of SARS-CoV-2—Escape from the NA extraction kit-shortage, Copenhagen, Denmark, march 2020. Eurosurveillance. 2020;25(14):2000398. doi:10.2807/1560-7917.ES.2020.25.14.2000398.
Forni and Mantovani, 2021 Forni G., Mantovani A. COVID-19 vaccines: Where we stand and challenges ahead. Cell Death and Differentiation. 2021;28(2):626–639. doi:10.1038/s41418-020-00720-9.
Ganbaatar and Liu, 2021 Ganbaatar U., Liu C. CRISPR-based COVID-19 testing: Toward next-generation point-of-care diagnostics. Frontiers in Cellular and Infection Microbiology. 2021;11:663949doi:10.3389/fcimb.2021.663949.
Greaney et al., 2021 Greaney A.J., Starr T.N., Barnes C.O., Weisblum Y., Schmidt F., Caskey M., et al. Mutational escape from the polyclonal antibody response to SARS-CoV-2 infection is largely shaped by a single class of antibodies [preprint]. Microbiology. 2021;doi:10.1101/2021.03.17.435863.
Green and Sambrook, 2018 Green M.R., Sambrook J. Quantification of RNA by real-time reverse transcription-polymerase chain reaction (RT-PCR). Cold Spring Harbor Protocols. 2018;2018(10):doi:10.1101/pdb.prot095042 pdb.prot095042.
Grieshaber, MacKenzie, Vörös and Reimhult, 2008 Grieshaber D., MacKenzie R., Vörös J., Reimhult E. Electrochemical biosensors—Sensor principles and architectures. Sensors (Basel, Switzerland). 2008;8(3):1400–1458. doi:10.3390/s80314000.
Guglielmi, 2020 Guglielmi G. Fast coronavirus tests: What they can and can't do. Nature. 2020;585(7826):496–498. doi:10.1038/d41586-020-02661-2.
Guo et al., 2020 Guo L., Sun X., Wang X., Liang C., Jiang H., Gao Q., et al. SARS-CoV-2 detection with CRISPR diagnostics. Cell Discovery. 2020;6(1):34. doi:10.1038/s41421-020-0174-y.
Harvey et al., 2021 Harvey W.T., Carabelli A.M., Jackson B., Gupta R.K., Thomson E.C., Harrison E.M., et al. SARS-CoV-2 variants, spike mutations and immune escape. Nature Reviews. Microbiology. 2021;19 (7):409–424. doi:10.1038/s41579-021-00573-0.
Heid, Stevens, Livak and Williams, 1996 Heid C.A., Stevens J., Livak K.J., Williams P.M. Real time quantitative PCR. Genome Research. 1996;6(10):986–994. doi:10.1101/gr.6.10.986.
Holder, 2021 Holder, J. (2021). Covid world vaccination tracker. The New York times.https://www.nytimes.com/interactive/2021/world/covid-vaccinations-tracker.html
Huang et al., 2021 Huang L., Ding L., Zhou J., Chen S., Chen F., Zhao C., et al. One-step rapid quantification of SARS-CoV-2 virus particles via low-cost nanoplasmonic sensors in generic microplate reader and point-of-care device. Biosensors & Bioelectronics. 2021;171:112685doi:10.1016/j.bios.2020.112685.
Hussain, Rupp, Wendel and Gehring, 2018 Hussain M., Rupp F., Wendel H.P., Gehring F.K. Bioapplications of acoustic crystals—A review. TrAC Trends in Analytical Chemistry. 2018;102:194–209. doi:10.1016/j.trac.2018.02.009.
Ishikawa et al., 2009 Ishikawa F.N., Chang H.-K., Curreli M., Liao H.-I., Olson C.A., Chen P.-C., et al. Label-free, electrical detection of the SARS virus N-protein with nanowire biosensors utilizing antibody mimics as capture probes. ACS Nano. 2009;3(5):1219–1224. doi:10.1021/nn900086c.
Jackson, Zhang, Farzan and Choe, 2021 Jackson C.B., Zhang L., Farzan M., Choe H. Functional importance of the D614G mutation in the SARS-CoV-2 spike protein. Biochemical and Biophysical Research Communications. 2021;538:108–115. doi:10.1016/j.bbrc.2020.11.026.
Kaminski, Abudayyeh, Gootenberg, Zhang and Collins, 2021 Kaminski M.M., Abudayyeh O.O., Gootenberg J.S., Zhang F., Collins J.J. CRISPR-based diagnostics. Nature Biomedical Engineering. 2021;5(7):643–656. doi:10.1038/s41551-021-00760-7.
Kellner, Koob, Gootenberg, Abudayyeh and Zhang, 2019 Kellner M.J., Koob J.G., Gootenberg J.S., Abudayyeh O.O., Zhang F. SHERLOCK: Nucleic acid detection with CRISPR nucleases. Nature Protocols. 2019;14(10):2986–3012. doi:10.1038/s41596-019-0210-2.
Khateeb, Li and Zhang, 2021 Khateeb J., Li Y., Zhang H. Emerging SARS-CoV-2 variants of concern and potential intervention approaches. Critical Care. 2021;25(1):244. doi:10.1186/s13054-021-03662-x.
Kim et al., 2020 Kim Y., Yaseen A.B., Kishi J.Y., Hong F., Saka S.K., Sheng K., et al. Single-strand RPA for rapid and sensitive detection of SARS-CoV-2 RNA. 2020.doi:10.1101/2020.08.17.20177006.
Kim et al., 2021 Kim H.-Y., Lee J.-H., Kim M.J., Park S.C., Choi M., Lee W., et al. Development of a SARS-CoV-2-specific biosensor for antigen detection using scFv-fc fusion proteins. Biosensors & Bioelectronics. 2021;175:112868doi:10.1016/j.bios.2020.112868.
Korber et al., 2020 Korber B., Fischer W.M., Gnanakaran S., Yoon H., Theiler J., Abfalterer W., et al. Tracking changes in SARS-CoV-2 spike: Evidence that D614G increases infectivity of the COVID-19 virus. Cell. 2020;182(4):doi:10.1016/j.cell.2020.06.043 812–827.e19.
Laffeber, de Koning, Kanaar and Lebbink, 2021 Laffeber C., de Koning K., Kanaar R., Lebbink J.H.G. Experimental evidence for enhanced receptor binding by rapidly spreading SARS-CoV-2 Variants. Journal of Molecular Biology. 2021;433(15):167058doi:10.1016/j.jmb.2021.167058.
Lee et al., 2021 Lee J.-H., Choi M., Jung Y., Lee S.K., Lee C.-S., Kim J., et al. A novel rapid detection for SARS-CoV-2 spike 1 antigens using human angiotensin converting enzyme 2 (ACE2). Biosensors & Bioelectronics. 2021;171:112715doi:10.1016/j.bios.2020.112715.
Li et al., 2020 Li Z., Yi Y., Luo X., Xiong N., Liu Y., Li S., et al. Development and clinical application of a rapid IgM-IgG combined antibody test for SARS-CoV-2 infection diagnosis. Journal of Medical Virology. 2020;92(9):1518–1524. doi:10.1002/jmv.25727.
Li et al., 2021 Li B., Deng A., Li K., Hu Y., Li Z., Xiong Q., et al. Viral infection and transmission in a large, well-traced outbreak caused by the SARS-CoV-2 Delta variant [preprint]. Epidemiology. 2021;doi:10.1101/2021.07.07.21260122.
Liu et al., 2021 Liu H., Dai E., Xiao R., Zhou Z., Zhang M., Bai Z., et al. Development of a SERS-based lateral flow immunoassay for rapid and ultra-sensitive detection of anti-SARS-CoV-2 IgM/IgG in clinical samples. Sensors and Actuators B: Chemical. 2021;329:129196doi:10.1016/j.snb.2020.129196.
Mahari, Roberts, Shahdeo and Gandhi, 2020 Mahari S., Roberts A., Shahdeo D., Gandhi S. ECovSens-ultrasensitive novel in-house built printed circuit board based electrochemical device for rapid detection of nCovid-19 antigen, a spike protein domain 1 of SARS-CoV-2. 2020.doi:10.1101/2020.04.24.059204.
Martin et al., 2020 Martin J., Klapsa D., Wilton T., Zambon M., Bentley E., Bujaki E., et al. Tracking SARS-CoV-2 in sewage: Evidence of changes in virus variant predominance during COVID-19 pandemic. Viruses. 2020;12(10):1144. doi:10.3390/v12101144.
Mavrikou, Moschopoulou, Tsekouras and Kintzios, 2020 Mavrikou S., Moschopoulou G., Tsekouras V., Kintzios S. Development of a portable, ultra-rapid and ultra-sensitive cell-based biosensor for the direct detection of the SARS-COV-2 S1 spike protein antigen. Sensors (Switzerland). 2020;20(11):doi:10.3390/s20113121.
McAulay et al., 2020 McAulay K., Bryan A., Greninger A.L., Grill F., Lake D., Kaleta E.J., et al. Retrospective clinical evaluation of 4 lateral flow assays for the detection of SARS-CoV-2 IgG. Diagnostic Microbiology and Infectious Disease. 2020;98(3):115161doi:10.1016/j.diagmicrobio.2020.115161.
Meng et al., 2021 Meng B., Kemp S.A., Papa G., Datir R., Ferreira I.A.T.M., Marelli S., et al. Recurrent emergence of SARS-CoV-2 spike deletion H69/V70 and its role in the Alpha variant B.1.1.7. Cell Reports. 2021;35 (13):109292. doi:10.1016/j.celrep.2021.109292.
Moitra, Alafeef, Dighe, Frieman and Pan, 2020 Moitra P., Alafeef M., Dighe K., Frieman M.B., Pan D. Selective naked-eye detection of SARS-CoV-2 mediated by N gene targeted antisense oligonucleotide capped Plasmonic nanoparticles. ACS Nano. 2020;14(6):7617–7627. doi:10.1021/acsnano.0c03822.
Morley, 2014 Morley A.A. Digital PCR: A brief history. Biomolecular Detection and Quantification. 2014;1(1):1–2. doi:10.1016/j.bdq.2014.06.001.
Morris et al., 2021 Morris C.P., Luo C.H., Amadi A., Schwartz M., Gallagher N., Ray S.C., et al. An update on SARS-CoV-2 diversity in the United States National Capital Region: Evolution of novel and Variants of concern. Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America. 2021;ciab636. https://doi.org/10.1093/cid/ciab636.
Motozono et al., 2021 Motozono C., Toyoda M., Zahradnik J., Ikeda T., Saito A., Tan T.S., et al. An emerging SARS-CoV-2 mutant evading cellular immunity and increasing viral infectivity [preprint]. Microbiology. 2021;doi:10.1101/2021.04.02.438288.
Nguyen et al., 2021 Nguyen P.Q., Soenksen L.R., Donghia N.M., Angenent-Mari N.M., de Puig H., Huang A., et al. Wearable materials with embedded synthetic biology sensors for biomolecule detection. Nature Biotechnology. 2021;1–9:doi:10.1038/s41587-021-00950-3.
Nolan, Hands and Bustin, 2006 Nolan T., Hands R.E., Bustin S.A. Quantification of mRNA using real-time RT-PCR. Nature Protocols. 2006;1(3):1559–1582. doi:10.1038/nprot.2006.236.
Ong et al., 2021 Ong S.W.X., Chiew C.J., Ang L.W., Mak T.-M., Cui L., Toh M.P.H.S., et al. Clinical and virological features of SARS-CoV-2 variants of concern: A retrospective cohort study comparing B.1.1.7 (Alpha), B.1.315 (Beta), and B.1.617.2 (Delta). Clinical Infectious Diseases: An Official Publication of the Infectious Diseases Society of America. 2021;ciab721doi:10.1093/cid/ciab721.
Palaz, Kalkan, Tozluyurt and Ozsoz, 2021 Palaz F., Kalkan A.K., Tozluyurt A., Ozsoz M. CRISPR-based tools: Alternative methods for the diagnosis of COVID-19. Clinical Biochemistry. 2021;89:1–13. doi:10.1016/j.clinbiochem.2020.12.011.
Pandey, 2020 Pandey L.M. Design of engineered surfaces for prospective detection of SARS-CoV-2 using quartz crystal microbalance-based techniques. Expert Review of Proteomics. 2020;17(6):425–432. doi:10.1080/14789450.2020.1794831.
Paweletz, Charboneau and Liotta, 2001 Paweletz C.P., Charboneau L., Liotta L.A. Micro RT-PCR. Current Protocols in Cell Biology. 2001;10(1):A.3G.1–A.3G.6. doi:10.1002/0471143030.cba03gs10.
Peacock et al., 2021 Peacock T.P., Goldhill D.H., Zhou J., Baillon L., Frise R., Swann O.C., et al. The furin cleavage site in the SARS-CoV-2 spike protein is required for transmission in ferrets. Nature Microbiology. 2021;6(7):899–909. doi:10.1038/s41564-021-00908-w.
Peñarrubia et al., 2020 Peñarrubia L., Ruiz M., Porco R., Rao S.N., Juanola-Falgarona M., Manissero D., et al. Multiple assays in a real-time RT-PCR SARS-CoV-2 panel can mitigate the risk of loss of sensitivity by new genomic variants during the COVID-19 outbreak. International Journal of Infectious Diseases. 2020;97:225–229. doi:10.1016/j.ijid.2020.06.027.
Planas et al., 2021 Planas D., Veyer D., Baidaliuk A., Staropoli I., Guivel-Benhassine F., Rajah M.M., et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature. 2021;596(7871):276–280. doi:10.1038/s41586-021-03777-9.
Qiu et al., 2020 Qiu G., Gai Z., Tao Y., Schmitt J., Kullak-Ublick G.A., Wang J. Dual-functional plasmonic photothermal biosensors for highly accurate severe acute respiratory syndrome coronavirus 2 detection. ACS Nano. 2020;14(5):5268–5277. doi:10.1021/acsnano.0c02439.
Rahimi et al., 2021 Rahimi H., Salehiabar M., Barsbay M., Ghaffarlou M., Kavetskyy T., Sharafi A., et al. CRISPR systems for COVID-19 diagnosis. ACS Sensors. 2021;6(4):1430–1445. doi:10.1021/acssensors.0c02312.
Ramos, Vela-Pérez, Ferrández, Kubik and Ivorra, 2021 Ramos A.M., Vela-Pérez M., Ferrández M.R., Kubik A.B., Ivorra B. Modeling the impact of SARS-CoV-2 variants and vaccines on the spread of COVID-19. Communications in Nonlinear Science and Numerical Simulation. 2021;102:105937doi:10.1016/j.cnsns.2021.105937.
Reardon, 2021 Reardon S. How the Delta variant achieves its ultrafast spread. Nature. 2021;doi:10.1038/d41586-021-01986-w.
Ruiz-Vega, Soler and Lechuga, 2021 Ruiz-Vega G., Soler M., Lechuga L.M. Nanophotonic biosensors for point-of-care COVID-19 diagnostics and coronavirus surveillance. Journal of Physics: Photonics. 2021;3(1):011002doi:10.1088/2515-7647/abd4ee.
Salian et al., 2021 Salian V.S., Wright J.A., Vedell P.T., Nair S., Li C., Kandimalla M., et al. COVID-19 transmission, current treatment, and future therapeutic strategies. Molecular Pharmaceutics. 2021;18(3):754–771. doi:10.1021/acs.molpharmaceut.0c00608.
Sallam and Mahafzah, 2021 Sallam M., Mahafzah A. Molecular analysis of SARS-CoV-2 genetic lineages in Jordan: Tracking the introduction and spread of COVID-19 UK variant of concern at a country level. Pathogens. 2021;10(3):302. doi:10.3390/pathogens10030302.
SARS-CoV-2 Total ab assay, n.d SARS-CoV-2 Total ab assay. (n.d.). Bio-Rad Laboratories. Retrieved August 25, 2021e, from https://www.bio-rad.com/en-us/product/sars-cov-2-total-ab-assay?ID=QADMT6E08O1Y
Seo et al., 2020 Seo G., Lee G., Kim M.J., Baek S.-H., Choi M., Ku K.B., et al. Rapid detection of COVID-19 causative virus (SARS-CoV-2) in human nasopharyngeal swab specimens using Field-effect transistor-based biosensor. ACS Nano. 2020;14(4):5135–5142. doi:10.1021/acsnano.0c02823.
Shahhosseini, Babuadze, Wong and Kobinger, 2021 Shahhosseini N., Babuadze G.G., Wong G., Kobinger G.P. Mutation signatures and in silico docking of novel SARS-CoV-2 variants of concern. Microorganisms. 2021;9(5):926. doi:10.3390/microorganisms9050926.
Shrivastav, Cvelbar and Abdulhalim, 2021 Shrivastav A.M., Cvelbar U., Abdulhalim I. A comprehensive review on plasmonic-based biosensors used in viral diagnostics. Communications Biology. 2021;4(1):70. doi:10.1038/s42003-020-01615-8.
Shu et al., 2021 Shu B., Kirby M.K., Davis W.G., Warnes C., Liddell J., Liu J., et al. Multiplex real-time reverse transcription PCR for influenza A virus, influenza B virus, and severe acute respiratory syndrome coronavirus 2. Emerging Infectious Diseases Journal—CDC. 2021. ;27(7). https://doi.org/10.3201/eid2707.210462.
Shukha, Makhoul, Abu-Elhija, Hayek and Hamoud, 2021 Shukha Y., Makhoul K., Abu-Elhija J., Hayek T., Hamoud S. A case of severe COVID-19 pneumonia diagnosed in bronchoscopy with negative repeated nasopharyngeal swabs. Journal of Medical Cases. 2021;12(6):217–219.
Smyrlaki et al., 2020 Smyrlaki I., Ekman M., Lentini A., Rufino de Sousa N., Papanicolaou N., Vondracek M., et al. Massive and rapid COVID-19 testing is feasible by extraction-free SARS-CoV-2 RT-PCR. Nature Communications. 2020;11(1):4812. doi:10.1038/s41467-020-18611-5.
Tegally et al., 2021 Tegally H., Wilkinson E., Giovanetti M., Iranzadeh A., Fonseca V., Giandhari J., et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature. 2021;592(7854):438–443. doi:10.1038/s41586-021-03402-9.
Tian, Gao, Fock, Dufva and Hansen, 2020 Tian B., Gao F., Fock J., Dufva M., Hansen M.F. Homogeneous circle-to-circle amplification for real-time optomagnetic detection of SARS-CoV-2 RdRp coding sequence. Biosensors & Bioelectronics. 2020;165:112356doi:10.1016/j.bios.2020.112356.
Uddin et al., 2020 Uddin M., Mustafa F., Rizvi T.A., Loney T., Al Suwaidi H., Al-Marzouqi A.H.H., et al. SARS-CoV-2/COVID-19: Viral genomics, epidemiology, vaccines, and therapeutic interventions. Viruses. 2020;12(5):526. doi:10.3390/v12050526.
Unser, Bruzas, He and Sagle, 2015 Unser S., Bruzas I., He J., Sagle L. Localized surface plasmon resonance biosensing: Current challenges and approaches. Sensors (Basel, Switzerland). 2015;15(7):15684–15716. doi:10.3390/s150715684.
van Dorp et al., 2020 van Dorp L., Acman M., Richard D., Shaw L.P., Ford C.E., Ormond L., et al. Emergence of genomic diversity and recurrent mutations in SARS-CoV-2. Infection, Genetics and Evolution. 2020;83:104351doi:10.1016/j.meegid.2020.104351.
Vandenberg, Martiny, Rochas, van Belkum and Kozlakidis, 2020 Vandenberg O., Martiny D., Rochas O., van Belkum A., Kozlakidis Z. Considerations for diagnostic COVID-19 tests. Nature Reviews. Microbiology. 2020;1–13. doi:10.1038/s41579-020-00461-z.
Vogels et al., 2021 Vogels C., Watkins A.E., Harden C.A., Brackney D.E., Shafer J., Wang J., et al. SalivaDirect: A simplified and flexible platform to enhance SARS-CoV-2 testing capacity. Med (New York, N.Y.). 2021;2(3):263–280.e6. doi:10.1016/j.medj.2020.12.010.
Volz et al., 2021 Volz E., Mishra S., Chand M., Barrett J.C., Johnson R., Geidelberg L., et al. Assessing transmissibility of SARS-CoV-2 lineage B.1.1.7 in England. Nature. 2021. ;593(7858):266–269. https://doi.org/10.1038/s41586-021-03470-x.
Wang et al., 2020 Wang C., Yang X., Gu B., Liu H., Zhou Z., Shi L., et al. Sensitive and simultaneous detection of SARS-CoV-2-specific IgM/IgG using lateral flow immunoassay based on dual-mode quantum dot nanobeads. Analytical Chemistry. 2020;92(23):15542–15549. doi:10.1021/acs.analchem.0c03484.
Wang et al., 2021 Wang P., Nair M.S., Liu L., Iketani S., Luo Y., Guo Y., et al. Antibody resistance of SARS-CoV-2 variants B.1.351 and B.1.1.7. Nature. 2021;593(7857):130–135. doi:10.1038/s41586-021-03398-2.
West, Gronvall and Kobokovich, 2021 West R., Gronvall G., Kobokovich A. Variants, vaccines and what they mean for COVID-19 testing. Johns Hopkins Bloomberg School of Public Health. 2021. https://www.jhsph.edu/covid-19/articles/variants-vaccines-and-what-they-mean-for-covid19-testing.html.
WHO COVID-19 Dashboard, 2020 WHO COVID-19 Dashboard. WHO COVID-19 dashboard. Geneva: World Health Organization ; 2020 Available online:Https://covid19.who.int/ (last cited: [Date]). https://covid19.who.int.
Wolk, Kaleta and Wysocki, 2012 Wolk D.M., Kaleta E.J., Wysocki V.H. PCR–electrospray ionization mass spectrometry. The Journal of Molecular Diagnostics: JMD. 2012;14(4):295–304. doi:10.1016/j.jmoldx.2012.02.005.
Wu and Ju, 2012 Wu J., Ju H.X. 3.07—Clinical immunoassays and immunosensing. In: Pawliszyn J., ed. Comprehensive sampling and sample preparation. Academic Press; 2012:143–167. doi:10.1016/B978-0-12-381373-2.00071-5.
Wu, Xu, Zhu and Xia, 2020 Wu Y., Xu W., Zhu Z., Xia X. Laboratory verification of an RT-PCR assay for SARS-CoV-2. Journal of Clinical Laboratory Analysis. 2020;34(10):e23507doi:10.1002/jcla.23507.
Xing et al., 2020 Xing W., Liu Y., Wang H., Li S., Lin Y., Chen L., et al. A high-throughput, multi-index isothermal amplification platform for rapid detection of 19 types of common respiratory viruses including SARS-CoV-2. Engineering (Beijing, China). 2020;6(10):1130–1140. doi:10.1016/j.eng.2020.07.015.
Yakoh et al., 2021 Yakoh A., Pimpitak U., Rengpipat S., Hirankarn N., Chailapakul O., Chaiyo S. Paper-based electrochemical biosensor for diagnosing COVID-19: Detection of SARS-CoV-2 antibodies and antigen. Biosensors & Bioelectronics. 2021;176:112912doi:10.1016/j.bios.2020.112912.
Yamamoto et al., 2021 Yamamoto S., Tanaka A., Kobayashi S., Oshiro Y., Ozeki M., Maeda K., et al. Consistency of the results of rapid serological tests for SARS-CoV-2 among healthcare workers in a large national hospital in Tokyo, Japan. Global Health & Medicine. 2021. ;3(2):90–94. https://doi.org/10.35772/ghm.2021.01022.
Yang and Du, 2021 Yang Y., Du L. SARS-CoV-2 spike protein: A key target for eliciting persistent neutralizing antibodies. Signal Transduction and Targeted Therapy. 2021;6(1):95. doi:10.1038/s41392-021-00523-5.
Zhang et al., 2020 Zhang W., Du R.-H., Li B., Zheng X.-S., Yang X.-L., Hu B., et al. Molecular and serological investigation of 2019-nCoV infected patients: Implication of multiple shedding routes. Emerging Microbes & Infections. 2020;9(1):386–389. doi:10.1080/22221751.2020.1729071.
Zhao et al., 2021 Zhao H., Liu F., Xie W., Zhou T.-C., OuYang J., Jin L., et al. Ultrasensitive supersandwich-type electrochemical sensor for SARS-CoV-2 from the infected COVID-19 patients using a smartphone. Sensors and Actuators. B, Chemical. 2021;327:128899doi:10.1016/j.snb.2020.128899.
Zhu et al., 2020 Zhu X., Wang X., Han L., Chen T., Wang L., Li H., et al. Multiplex reverse transcription loop-mediated isothermal amplification combined with nanoparticle-based lateral flow biosensor for the diagnosis of COVID-19. Biosensors & Bioelectronics. 2020;166:112437doi:10.1016/j.bios.2020.112437.
Further reading
Access SARS-CoV-2 IgM Antibody Test. (n.d.). Beckman Coulter. Retrieved August 25, 2021, from https://www.beckmancoulter.com/products/immunoassay/access-sars-cov-2-igm-antibody-test
ACCESS SARS-CoV-2 IgM. (2020). Beckman Coulter. https://www.fda.gov/media/142911/download
Coronavirus (COVID-19) Update: FDA Authorizes First Test that Detects Neutralizing Antibodies from Recent or Prior SARS-CoV-2 Infection. FDA; FDA. https://www.fda.gov/news-events/press-announcements/coronavirus-covid-19-update-fda-authorizes-first-test-detects-neutralizing-antibodies-recent-or. 2020, November 6.
Diazyme DZ-Lite SARS-CoV-2 IgM CLIA Kit. (n.d.). Diazyme Laboratories, Inc. Retrieved August 25, 2021, from https://www.diazyme.com/covid-19-antibody-tests/dz-lite-sars-cov-2-igm-clia-kit
Diazyme Laboratories, Inc. DIAZYME DZ-LITE SARS-CoV-2 IgM CLIA KIT. Diazyme. 2020 August 14;1–4. https://www.fda.gov/media/141255/download.
Elecsys® anti-SARS-CoV-2. (n.d.). Diagnostics. Retrieved August 24, 2021b, from https://diagnostics.roche.com/us/en/products/params/elecsys-anti-sars-cov-2.html
ELISA: Purpose, Procedure, and Results. (2012, July 19). Healthline. https://www.healthline.com/health/elisa
In Vitro Diagnostics EUAs—Molecular Diagnostic Tests for SARS-CoV-2. (2021, August 30). U.S. Food & Drug Administration; FDA. https://www.fda.gov/medical-devices/coronavirus-disease-2019-covid-19-emergency-use-authorizations-medical-devices/in-vitro-diagnostics-euas-molecular-diagnostic-tests-sars-cov-2
Innovita 2019-nCoV Ab Test (Colloidal Gold)—Instructions for Use. (n.d.). https://www.fda.gov/media/144071/download
Labcorp's COVID-19 RT-PCR Test EUA Summary. (2021). 30. https://www.fda.gov/media/136151/download
OmniPATH COVID-19 Total Antibody ELISA Test | COVID-19 Antibody Testing. (n.d.). Thermo Fisher Scientific. Retrieved August 25, 2021, from https://www.thermofisher.com/covid-19-antibody-testing/us/en/solutions/OmniPATH-COVID19-Total-Antibody-ELISA-Test.html
OmniPATH™ COVID-19 Total Antibody ELISA Test. (n.d.). https://www.fda.gov/media/142700/download
Platelia SARS-CoV-2 Total Ab. (n.d.). https://www.fda.gov/media/137493/download
Roche develops new serology test to detect COVID-19 antibodies. (n.d.). Diagnostics. Retrieved August 24, 2021d, from shttps://diagnostics.roche.com/global/en/news-listing/2020/roche-develops-new-serology-test-to-detect-covid-19-antibodies.html
SalivaDirect Papers & Protocols. (n.d.). Retrieved August 31, 2021, from https://ysph.yale.edu/salivadirect/publications/
SARS-CoV-2 Total Antibody assay (CV2T). (n.d.). https://www.fda.gov/media/138757/download
Serology: Purpose, Results, and Aftercare. (2012, June 8). Healthline. https://www.healthline.com/health/serology
TaqPath COVID-19 multiplex diagnostic solution—US. (n.d.). Retrieved August 31, 2021, from //www.thermofisher.com/us/en/home/clinical/clinical-genomics/pathogen-detection-solutions/covid-19-sars-cov-2/multiplex.html
TaqPath™ COVID-19 Combo Kit and TaqPath™ COVID-19 Combo Kit Advanced INSTRUCTIONS FOR USE. (n.d.). https://www.fda.gov/media/136112/download
Testing Tool Kit, 2021. https://www.centerforhealthsecurity.org/covid-19TestingToolkit/testing-basics/types-of-COVID-19-tests/serology-tests.html
Variants, Vaccines and What They Mean For COVID-19 Testing | Johns Hopkins Bloomberg School of Public Health. (2021, February 2). Johns Hopkins Bloomberg School of Public Health. https://publichealth.jhu.edu/2021/variants-vaccines-and-what-they-mean-for-covid-19-testing
What is Chemiluminescent immunoassay? | Lorne Laboratories UK. (n.d.). Lorne Laboratories Ltd. Retrieved August 30, 2021, from https://www.lornelabs.com/news-events/blog/what-is-chemiluminescent-immunoassay