Chinekwu Sherridan Nwagwua,*; Chinenye Nnenna Ugwub; John Dike Nwabueze Ogbonnaa; Adaeze Linda Onugwua; Chinazom Precious Agboa; Adaeze Chidiebere Echezonaa; Ezinwanne Nneoma Ezeibeb; Samuel Uzondua; Frankline Chimaobi Kenechukwua; Paul Achile Akpaa; Mumuni Audu Momoha; Petra Obioma Nnamania; Clemence Tariraic; Kenneth Chibuzor Ofokansia; Anthony Amaechi Attamaa a Drug Delivery & Nanomedicines Research Laboratory, Department of Pharmaceutics, University of Nigeria, Nsukka, Enugu State, Nigeria
b Department of Pharmaceutical Microbiology and Biotechnology, University of Nigeria, Nsukka, Enugu state, Nigeria
c Department of Pharmaceutical Sciences, Tshwane University of Technology, Pretoria, South Africa
* Corresponding author: email address: chinekwu.nwobi@unn.edu.ng
Abstract
The outbreak of the COVID-19 pandemic in 2019 has been one of the greatest challenges modern medicine and science has ever faced. It has affected millions of people around the world and altered human life and activities as we once knew. The high prevalence as well as an extended period of incubations which usually does not present with symptoms have played a formidable role in the transmission and infection of millions. A lot of research has been carried out on developing suitable treatment and effective preventive measures for the control of the pandemic. Preventive strategies which include social distancing, use of masks, washing of hands, and contact tracing have been effective in slowing the spread of the virus; however, the infectious nature of the SARS-COV-2 has made these strategies unable to eradicate its spread. In addition, the continuous increase in the number of cases and death, as well as the appearance of several variants of the virus, has necessitated the development of effective and safe vaccines in a bid to ensure that human activities can return to normalcy. Nanotechnology has been of great benefit in the design of vaccines as nano-sized materials have been known to aid the safe and effective delivery of antigens as well as serve as suitable adjuvants to potentiate responses to vaccines. There are only four vaccine candidates currently approved for use in humans while many other candidates are at various levels of development. This review seeks to provide updated information on the current nano-technological strategies employed in the development of COVID-19 vaccines.
Keywords
SARS-COV-2; COVID-19; RNA; Immunity; COVID-19 vaccines; Nanotechnology; Nanocarriers; Nanoparticles vaccine adjuvants
1: Introduction
The coronavirus disease 2019 (COVID-19) is caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and is one of the most difficult health crises that humanity has faced in recent years. The pandemic has affected millions of people across the globe causing harm to humans as well as the economies of nations. Several public health strategies such as the use of masks, social distancing, regular washing of hands as well as contact tracing, have been employed since the beginning of the outbreak to curtail the spread of the virus. However, these practices have not been able to completely prevent the widespread of the pandemic (Young, Thone, & Jik, 2021). Despite the tireless efforts of researchers and scientists all over the world, there is as of now, still, no cure for COVID-19, although the United States Food and Drugs Administration (FDA) recently approved the use of remdesivir for treatment, especially in severe cases of viral infection (Campos et al., 2020). The outbreak of the pandemic has stretched the limits of healthcare systems and challenged the management of the situation using conventional tools in the development of treatment therapies, vaccines, and other preventive as well as diagnostic measures (Campos et al., 2020). Vaccination over the years has proven to be the most effective approach for the control and prevention of infectious diseases. Therefore, the control and subsequent total eradication of the pandemic requires the development of effective and safe vaccine protocols (Chung, Beiss, Fiering, & Steinmetz, 2020). The major focus in the development of COVID-19 vaccines is to limit and possibly totally prevent severe illness as well as the widespread of the virus. Studies have suggested that neutralizing antibodies confer protection as they inhibit viral entry by preventing the interaction between the rapid binding domain (RBD) of the SARS-COV-2 spike (S) protein with the angiotensin-converting enzyme (ACE) 2 receptors. Other approaches have also explored the role of T-cells in clearing infected cells. Some other approaches suggest the best approach for developing an efficient as well as safe COVID-19 vaccine is to ensure a balance between antibody and T-cell responses (Chauhan et al., 2020). There are about 117 vaccine candidates in clinical development and 194 candidates in pre-clinical development, with only about four candidates approved for use in humans (World Health Organization, 2021). Nanotechnological interventions have provided immense benefits in the area of vaccine development over the years, from providing sustainable nanomaterials for the delivery of antigens to being utilized as vaccine adjuvants to potentiate immune responses to vaccines. This is particularly important for the development of COVID-19 vaccines as some studies suggest that the SARS-COV-2 is a functional nanomaterial, possessing nano-metric dimensions with a nano-core-shell structure (Ruiz-Hitzky et al., 2020). This study focuses on reviewing the contributions of nanotechnological approaches in the development of COVID-19 vaccines.
2: The structure and infection mechanism of SARS-COV-2
SARS-CoV-2 is a beta-coronavirus and a single-stranded positive-sense RNA (+ ssRNA) which is bigger than any other RNA virus in the genome of CoVs (27–32 kb) (Wang, Hu, et al., 2020; Wang, Peng, Xu, Cui, & Williams, 2020; Wang, Zhao, et al., 2020). The capsid outside the genome is created by the nucleocapsid protein (N), and the genome is further packed by an envelope made up of three structural proteins: membrane protein (M), spike protein (S), and envelope protein (E) (Wan, Shang, Graham, Baric, & Li, 2020). SARS-CoV-2 is encoded by four structural proteins (S, E, M, and N) and sixteen non-structural proteins (nsp1–16). The M protein is the most abundant which shapes the virions (envelop) and is comprised of three transmembrane territories, that encourage membrane curving and helps fix the nucleocapsid (Ahmed et al., 2021). On the other hand, protein E is the smallest structural protein and performs important roles in the assembly and discharge of viruses (DeDiego et al., 2007; Nieto-Torres et al., 2014). The N protein is associated with RNA which forms an envelope containing the nucleocapsid. Additionally, N protein is largely involved in the viral genome processes and replication cycle which involves the assembly, budding, and also stimulating the host cellular response to viral infection (Chang et al., 2006; Hurst, Koetzner, & Masters, 2009). The S proteins have polymers that get embedded in the virion, obtaining the crown-like appearance, which helps the virus connect to the host cell's surface receptors and combines the viral and host cell membranes, allowing the virus to enter the host cell (Boopathi, Poma, & Kolandaivel, 2020). The spike protein (S-protein) facilitates the coronavirus access into host cells (Wang, Hu, et al., 2020; Wang, Peng, et al., 2020; Wang, Zhao, et al., 2020) by attaching to the ACE-2 receptor which has high expression on some cell types, such as endothelial cells, alveolar cells, kidney cells, intestinal epithelial cells, monocytes/macrophages, in addition to neuro-epithelial cells and neurons (Wan et al., 2020; Zhou et al., 2020) and low-to-no expression, in most cells and organs of the immune system (blood cells, spleen, bone marrow, and blood vessels) (Garciá, Mancilla-Galindo, Paredes-Paredes, Tiburcio, & Ávila-Vanzzini, 2021). After the spike (S) protein binds to the ACE-2 receptor, cleavage by transmembrane proteases serine 2, cathepsin, or furin causes endocytosis and translocation of the SARS-CoV-2 into endosomes (Reza-Zaldıvar, 2021), or direct viral envelope fusion with host cell membrane for cell entrance (Sanclemente-Alaman et al., 2020). Since the SARS-CoV-2 S protein is highly glycosylated and remains mostly in a closed prefusion conformation (Turoňová et al., 2020), pre-activation of the S protein by furin protease is thought to be an essential step to expose its receptor-binding domain (Garciá et al., 2021).
3: Pathogenesis and clinical presentation of COVID-19
SARS-CoV-2 is the cause of COVID-19 disease. Three phases of SARS-CoV-2 infection match different clinical stages of the COVID-19 disease, based on the cells affected (Mason, 2020). The first phase is the asymptomatic state, at this point, the inhaled SARS-CoV-2 virus binds to epithelial cells in the nasal cavity and starts replicating as seen in Fig. 1. The presence of the virus can be confirmed by swabbing the nasal cavity (Mason, 2020). The viral load at this stage although very low is very infectious and presents a limited innate immune response. At this point, the virus can be transmitted even before the symptoms appear (Yang, 2021). The second phase is when the virus replicates and moves down the respiratory tract. This triggers an increase in the innate immune response. The virus and early markers of the innate immune system can be detected by nasal swabs or sputum. At this phase, there is a mild clinical manifestation of the COVID-19. Individuals affected at this point can be managed at home with conventional treatment of the patients' symptoms. (Wu & McGoogan, 2020). Only about 20% of the infected patients progresses to the final stage of the disease which presents with massive pulmonary infiltration (Wu & McGoogan, 2020). Here, the virus invades the gas exchange components of the lungs and contaminates the alveolar type II cells (Mason, 2020). SARS-CoV-2 reproduces within the T-cells, discharging a huge amount of viral particles which instigates the cells to encounter caspase-mediated cell death and die (Qian et al., 2012). This stage initiates severe flu-like symptoms that can progress to acute respiratory distress (ARD), pneumonia, renal failure as well as death (Ksiazek et al., 2003; Wang, Hu, et al., 2020; Wang, Peng, et al., 2020; Wang, Zhao, et al., 2020). Initial studies about COVID-I9 reported the effect of the disease mainly on the respiratory tract but also on the gastrointestinal tract (Fig. 1). However, more recent reports strongly suggest that the disease can lead to several extra-pulmonary complications and even multiple organ injuries. There have been reports of myocardial inflammation and venous thromboembolism especially with patients with pre-existing cardiovascular conditions (Sarkesh et al., 2020). Hematological complications such as lymphopenia, increased level of interleukin 6 (IL-6) and some cytokines, thrombocytopenia, neutrophilia caused by hyper-inflammatory and cytokine storm resulting in severe organ malfunction and physiologic decompensation have been reported in patients with severe symptoms of COVID-19 (Alexandrova, Beykov, Vassilev, Jukić, & Podlipnik, 2021). Viral infections have over the years been known to cause serious structural and functional damage to the nervous system (Rahman et al., 2020; Sarkesh et al., 2020). Several studies have reported some central nervous system (CNS) complications such as cerebral infarctions in patients with severe cases of COVID-19 disease (Macera, De Angelis, Sagnelli, & Coppola, 2020). Also, older patients particularly those with underlying ailments, are at high risk of impaired consciousness or delirium at the onset of acute infections. These patients also display symptoms like encephalopathy and confusion. Rare, nonspecific neurological symptoms have been reported in COVID-19 patients, including dizziness, headache (Fig. 1), muscle injury leading to myalgia, neuralgia, and ataxia (Harrison, Lin, & Wang, 2020). Several SARS COV2 infected patients have been reported to present with several muscular disorders, some of which include myopathy, acute quadriplegic myopathy, thick filament myopathy, and necrotizing myopathy (Gálvez-Barrón et al., 2021; Mba, Sharndama, Osondu-chuka, & Okeke, 2021). Although the aged population is particularly vulnerable to COVID-19, due to their weakened immune system, poor ability to heal damaged epithelium, and a reduced mucociliary clearance, which permits the virus to easily move to the lung's gas exchange components, children and young adults can also be infected by the virus (Chao et al., 2020; Ho et al., 2001) (Fig. 1).
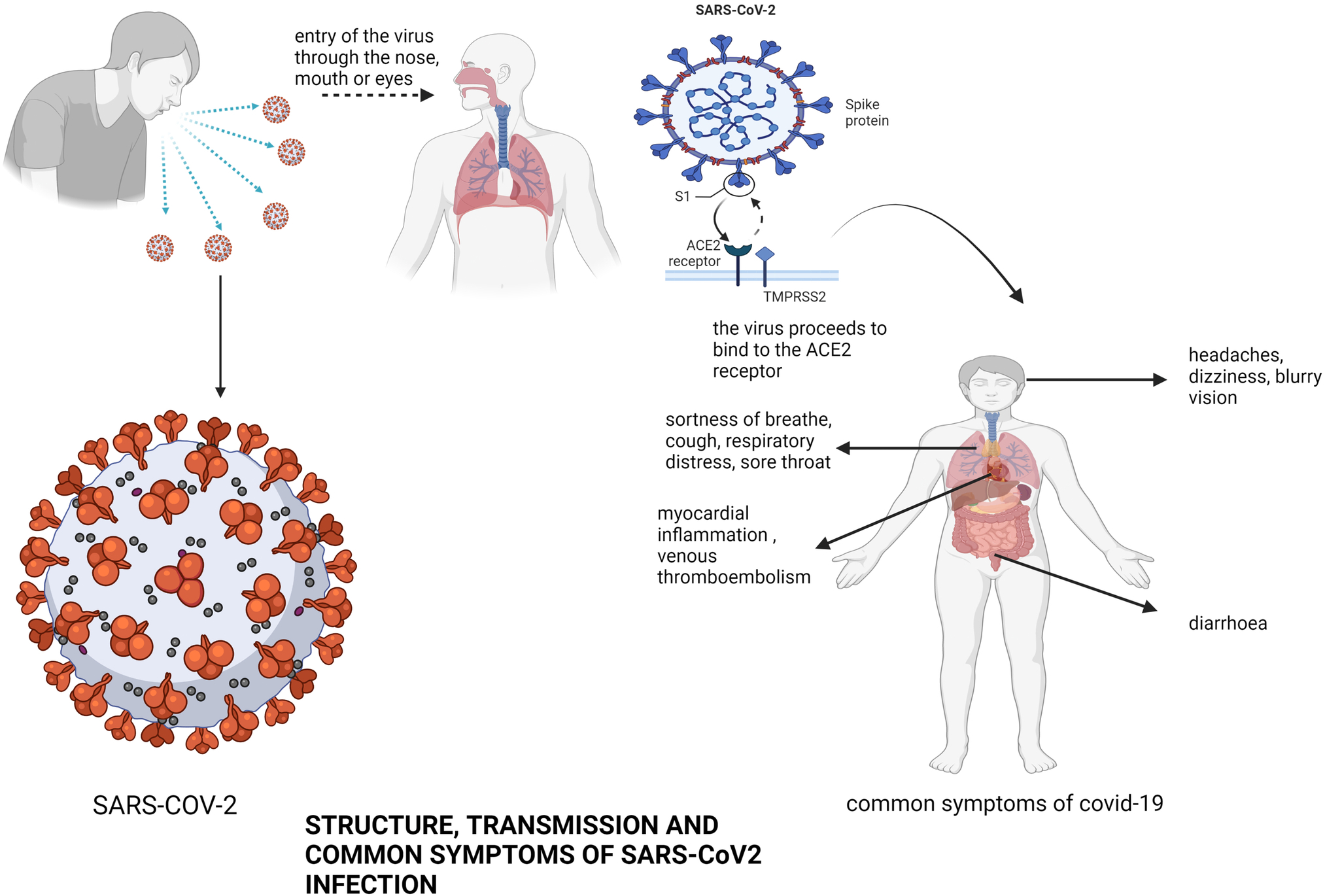 Fig. 1 Structure, transmission and common symptoms of SARS-CoV2 infection. Shown is the structure of SARS-CoV2, the common routes of transmission, and the common symptoms of infection with the virus).
Fig. 1 Structure, transmission and common symptoms of SARS-CoV2 infection. Shown is the structure of SARS-CoV2, the common routes of transmission, and the common symptoms of infection with the virus).
4: Vaccine development strategies and platforms
The use of a whole virus or bacterium components which trigger the immune system, or genetic materials that give instructions for the activation of the immune response is the basic approach to vaccine formulation. The vaccine strategy must be both safe and effective in producing strong and long-lasting protective immunity against the infectious pathogen, ideally with only one dose. Traditional vaccine development tactics are gradually giving way to newer and more advanced strategies such as viral vectored and nucleic acid vaccines, which are expanding the options in vaccine development. Vaccine platforms are divided into six main categories: live attenuated virus, inactivated or killed virus vaccine, protein subunit vaccines, virus-like particles (VLPs), viral-vectored vaccines, and nucleic acid-based (DNA or mRNA) vaccines. Fig. 2 provides a summary of the different vaccine strategies explored in the development of COVID-19 vaccines. Each of these approaches has its own set of benefits and drawbacks.
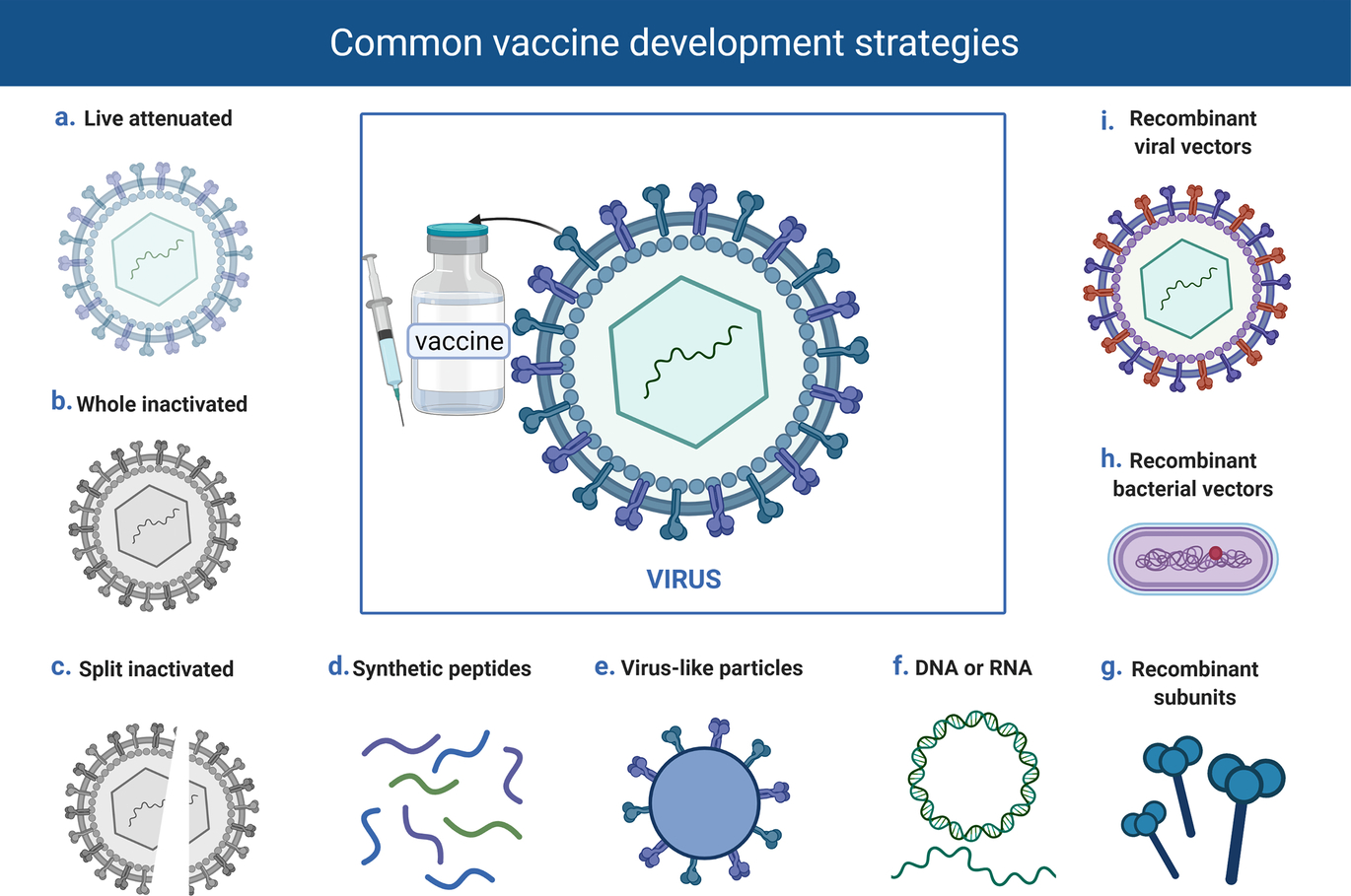 Fig. 2 Common vaccine strategies. This is a diagrammatic representation of the common vaccine strategies been explored in the development of COVD-19 vaccines.
Fig. 2 Common vaccine strategies. This is a diagrammatic representation of the common vaccine strategies been explored in the development of COVD-19 vaccines.
There are two major components of vaccines among others that should be given great consideration during the production process. Firstly, protein or polysaccharide antigens, which are supplied to or produced by the vaccine recipient. Another important component is an infection signal that alerts and activates the host immune system. Live attenuated vaccines can naturally provide both the antigen and the signal, while non-viral vaccine platforms can supply the antigens but frequently require adjuvant that gives the danger signals to the immune system. Fig. 3 further illustrates other basic components of a vaccine formulation.
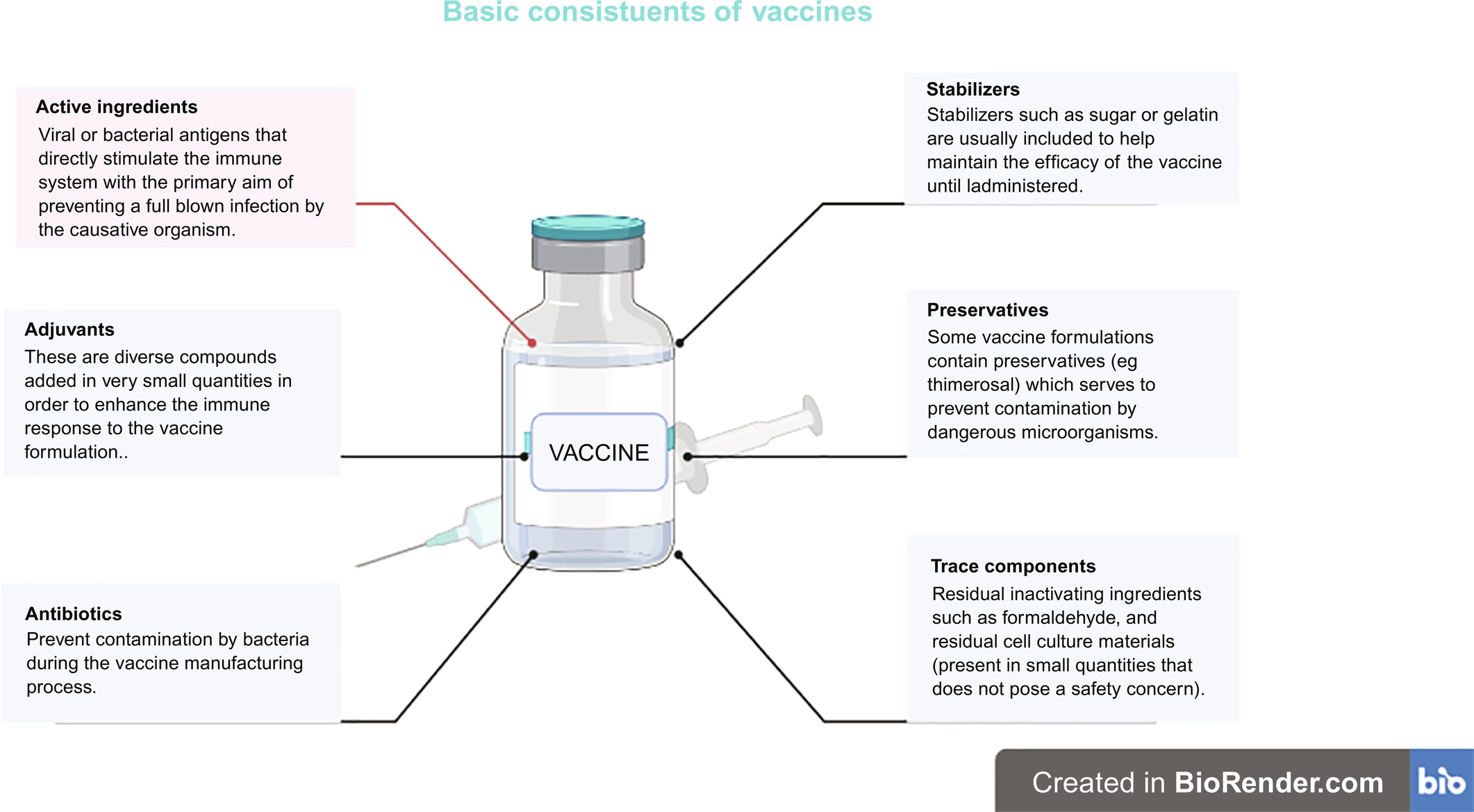 Fig. 3 Basic constituents of vaccines.
Fig. 3 Basic constituents of vaccines.
4.1: Live attenuated viral vaccines
Human vaccines based on a live attenuated vaccine platform which have been used successfully for decades include measles, mumps, rubella, poliomyelitis, and the bacillus Calmette–Guérin (BCG) vaccines (Plotkin, Orenstein, Offit, & Edwards, 2017). They are live, reproducing, but avirulent viruses or bacteria. This vaccine strategy is based on administering weakened but live strains of the actual pathogen that mimics the natural infection without causing disease in healthy individuals.
Attenuation of the pathogenic strains is traditionally achieved through in vitro passage that leads to mutation or loss of virulence genes. A new technique to design live attenuated virus strains is genetic modification via mutation or deletion of virulence genes. The mutants lose their pathogenicity property in the host cells but can replicate to a limited extent. Several genes possessed by coronaviruses are not required for replication and these genes can be deleted, leading to attenuation in vivo. Various non-structural proteins and structural E proteins can be deleted to yield vaccine strains of several coronaviruses. Deletion of the E protein leads to attenuation and generation of an efficacious vaccine strain (Almazán et al., 2013; Netland et al., 2010). There can be a problem of reversion of the attenuated strain to the virulent strain (Jimenez-Guardeño et al., 2015). Therefore, the deletion of virulence genes may provide a more effective mechanism of attenuation.
Codon deoptimization is another efficient approach to viral attenuation. The nucleic acid sequence is modified to use suboptimal codon pairs to encode the wild-type amino acid sequence, which considerably slows the translation of the viral protein during infection. This approach can produce a replication-competent but highly attenuated strain in vivo (Mueller et al., 2020). The recoded virus has an antigenic property similar to that of its pathogenic parents. Consequently, the attenuated viruses induce immune responses that are identical to those of virulent strains (Mueller et al., 2020).
The inability of the attenuated strain to revert genetically to become pathogenic is an important consideration in the generation of a live attenuated vaccine. Coronaviruses are known to often recombine in nature and this makes the development of an attenuated live vaccine against SARS-CoV-2 challenging. The attenuated strain could recombine with other wild coronaviruses resulting in a fully virulent strain. One of the drawbacks of live attenuated viruses is the use of exhaustively long cell or animal cultures in their development. Also, pre-existing cross-reactive immunity resulting from natural exposure with other human coronaviruses could limit the efficacy of SARS-CoV-2 vaccines developed using this platform. Another disadvantage of this technique is that attenuated vaccines cannot be given to immune-compromised persons since the attenuated agent would find a niche to multiply uncontrollably and, on rare occasions, revert to a wild-type phenotype, resulting in severe disease. As a result of these drawbacks, only two live attenuated SARS-CoV-2 vaccine candidates have reached clinical trials as seen in Table 1 (World Health Organization, 2021).
Table 1
|
Some vaccine candidates in clinical trials. |
||||
|
S/N |
Name of Vaccine |
Vaccine Platform |
Manufacturer |
Phase of clinical trial |
|
1. |
CoronaVac®:Inactivated SARSCoV-2 vaccine (Vero Cell) |
Inactivated, produced in vero cells |
Sinovac Research and Develop company Ltd |
Phase 4 |
|
2. |
Inactivated SARSCoV-2 vaccine (Vero Cell). BBIBP COVID-19 vaccine |
Inactivated, produced in Vero cells |
Sinopharm/Beijing Institute of Biological Products Co., Ltd. (BIBP) |
Phase 4 |
|
3. |
Coviran-Barkat® |
Inactivated |
Shifa Pharmed Industrial Co |
Phase 2/3 |
|
4. |
Inactivated SARS-CoV-2 Vaccine (Vero Cell) |
Inactivated, produced in Vero cells |
Sinopharm/Wuhan Institute of Biological Products Co., Ltd. (WIBP) |
Phase 4 |
|
5. |
ChAdOx1-S—(AZD1222) |
Viral vector (Non-replicating) |
AstraZeneca and University of Oxford |
Phase 4 |
|
6. |
Ad5-nCoV |
Viral vector (Non-replicating) |
CanSino Biological Inc./Beijing Institute of Biotechnology |
Phase 4 |
|
7. |
Gam-COVID-Vac Adeno-based (rAd26-S + rAd5-S) |
Viral vector (Non-replicating) |
Gamaleya Research Institute; Health Ministry of the Russian Federation |
Phase 3 |
|
8. |
Ad26.COV2·S |
Viral vector (Non-replicating) |
Janssen Pharmaceutical |
Phase 4 |
|
9. |
SARS-CoV-2 rS/Matrix M1-Adjuvant (Full length recombinant SARS CoV-2 |
Protein subunit |
Novavax |
Phase 3 |
|
10. |
mRNA-1273 |
RNA based vaccine |
Moderna + National Institute of Allergy and Infectious Diseases (NIAID) |
Phase 4 |
|
11. |
BNT162b2 (3 LNP-mRNAs), also known as “Comirnaty” |
RNA based vaccine |
Pfizer/BioNTech + Fosun Pharma |
Phase 4 |
|
12. |
Recombinant SARS-CoV-2 vaccine (CHO Cell) |
Protein subunit |
Anhui Zhifei Longcom Biopharmaceutical + Institute of Microbiology, Chinese Academy of Sciences |
Phase 3 |
|
13. |
CVnCoV Vaccine |
RNA based vaccine |
CureVac AG |
Phase 3 |
|
14. |
SARS-CoV-2 Vaccine, Inactivated (Vero Cell)/COVAXIN® |
Whole-Virion Inactivated |
Bharat Biotech International Ltd |
Phase 3 |
|
15. |
SARS-CoV-2 Vaccine, Inactivated (Vero Cell) |
Inactivated |
Institute of Medical Biotechnology Chinese Academy of Medical Sciences (IMBCAMS), China |
Phase 3 |
|
16. |
INO-4800 + electroporation |
DNA based vaccine |
Inovio Pharmaceuticals + International Vaccine Institute + Advaccine (Suzhou) Biopharmaceutical Co., Ltd |
Phase 3 |
|
17. |
AG0301-COVID19 |
DNA based vaccine |
AnGes + Takara Bio + Osaka University |
Phase 2/3 |
|
18. |
nCov vaccine |
DNA based vaccine |
Zydus Cadila |
Phase 3 |
|
19. |
GX-19 N |
DNA based vaccine |
Genexine Consortium |
Phase 2/3 |
|
20. |
KBP-COVID-19 (RBD-based) |
Protein subunit |
Kentucky Bioprocessing Inc. |
Phase 1/2 |
|
21. |
VAT00008: SARS-CoV-2 S protein with adjuvant |
Protein subunit |
Sanofi Pasteur + GSK |
Phase 3 |
|
22. |
ARCT-021 |
RNA based vaccine |
Arcturus Therapeutics |
Phase 2 |
|
23. |
RBD SARS-CoV-2 HBsAg VLP vaccine |
Virus like particle |
Serum Institute of India + Accelagen Pty + SpyBiotech |
Phase 1/2 |
|
24. |
GRAd-COV2 (Replication defective Simian Adenovirus (GRAd) encoding S) |
Viral vector (Non-replicating) |
ReiThera + Leukocare + Univercells |
Phase 2/3 |
|
25. |
VXA-CoV2–1 Ad5 adjuvanted Oral Vaccine platform |
Viral vector (Non-replicating) |
Vaxart |
Phase 2 |
|
26. |
MVA-SARS-2-S |
Viral vector (Non-replicating) |
University of Munich (Ludwig-Maximilians) |
Phase 1 |
|
27. |
Coronavirus-Like Particle COVID-19 (CoVLP) |
Virus like particle |
Medicago Inc. |
Phase 3 |
|
28. |
COVI-VAC |
Live attenuated virus |
Codagenix/Serum Institute of India |
Phase 3 |
|
29. |
Dendritic cell vaccine AV-COVID-19. A vaccine consisting of autologous dendritic cells loaded |
Viral vector (Replicating) + APC |
"Aivita Biomedical, Inc.; |
Phase 2 |
|
30. |
Inactivated COVID-19 vaccine |
Inactivated + alum |
KM Biologic Co Ltd |
Phase 2/3 |
|
31. |
Osvid-19® |
Inactivated |
Osve Pharmaceutical Company |
Phase 1 |
|
32. |
TURKOVAC® |
Inactivated |
Erciyes University and The Health Institute of Turkey (TUSEB) |
Phase 3 |
|
33. |
VB10.2210, DNA plasmid vaccine, encodes multiple immunogenic and conserved T cell epitopes spanning multiple antigens across the SARS-CoV-2 genome |
DNA based vaccine |
Vaccibody AS |
Phase 1/2 |
|
34. |
CoviVax® |
Inactivated whole virus |
National Research Centre, Egypt |
Phase 1 |
|
35. |
Adjuvanted inactivated vaccine against SARS-CoV 2 |
Inactivated |
The Scientific and Technological Research Council of Turkey (TUBITAK) |
Phase 1 |
|
36. |
Inactivated (NDV-based) Chimeric vaccine with or without adjuvant CpG 1018 |
Inactivated + CpG 1018 |
The Government Pharmaceutical Organization (GPO): PATH, Dynavax |
Phase 1/2 |
|
37. |
VLA2001® |
Inactivated |
Valneva, National Institution of Health Research UK |
Phase 3 |
|
38. |
QazCOVID-in® |
Inactivated |
Research Institute for Biological Safety Problems, Rep of Kazakhstan |
Phase 3 |
|
39. |
Kocak inactivated adjvant COVID-19 vaccine |
inactivated |
Kocak Farma |
Phase 1 |
|
40. |
CoviVac® |
inactivated |
Chumakov Federal Scientific Center for Development of Immune-and-Biological products |
Phase 1/2 |
|
41. |
MV-014-212, a live attenuated vaccine that expresses the spike (S) protein of SARS-CoV-2 |
Live attenuated virus |
Meissa Vaccines, Inc. |
Phase 1 |
|
42. |
VBI-2902a. An enveloped virus-like particle (eVLP) of SARS-CoV-2 spike (S) glycoprotein and aluminum phosphate adjuvant. |
Virus like particle |
VBI Vaccines Inc. |
Phase 1/2 |
|
43. |
SARS-CoV-2 VLP Vaccine |
Virus like particle |
The Scientific and Technological Research Council of Turkey |
Phase 2 |
|
44. |
ABNCoV2 capsid virus-like particle (cVLP) +/− adjuvant MF59 |
Virus like particle |
Radboud University |
Phase 1 |
4.2: Inactivated pathogen vaccines
Viruses inactivated through physical and chemical means have been used successfully in human vaccines against hepatitis A, polio, and influenza (Murdin, Barreto, & Plotkin, 1996; Vellozzi et al., 2009). In this platform, a dead form of the pathogen is used, thus ensuring a better safety profile than live attenuated vaccines. During the inactivation process using chemicals, heat, or radiation, some of these vaccine strains lose their immunogenicity making this platform less efficient than live attenuated pathogen immunization. Moreover, inactivated pathogen vaccines are poor inducers of cytotoxic CD8 + T cells, which are necessary for an effective COVID-19 vaccine. Inactivated viral vaccines require an adjuvant which are compounds that enhance and amplify the immune responses to the presence of an antigen. Inactivated viral vaccine also requires the administration of more than one dose to be effective. There are sixteen inactivated SARS-CoV-2 vaccines under clinical trials as seen in Table 1 (World Health Organization, 2021).
4.3: Protein subunit vaccines
Protein subunit vaccines are made up of specific isolated proteins from pathogenic bacteria or viruses. The advances in laboratory techniques that ushered in the era of genetic engineering in the 1970s resulted in the first recombinant protein vaccine, the hepatitis B vaccine (Vellozzi et al., 2009). Protein subunit vaccines are generated through recombinant technology or protein isolation and purification methods after cultivating large amounts of the pathogen. Instead of the whole pathogen, a subunit of it that stimulates the immune system is used. Proteins or peptides alone do not elicit a strong immunogenic response and thus require both adjuvant and booster doses. Just like an inactivated viral vaccine, subunit vaccines are poor inducers of CD8 + T cell responses. This platform eliminates the possibility of severe adverse effects, but there is a need for repeated administration and incorporation of an adjuvant to create robust and long-lasting immunity. Antigen-presenting cells (APCs), activated by the adjuvant, take up the antigen when administered and present it to adaptive immune cells (Vellozzi et al., 2009). There are thirty-six COVID-19 subunit vaccines currently in clinical trials (Fig. 4), making this the most common platform (World Health Organization, 2021).
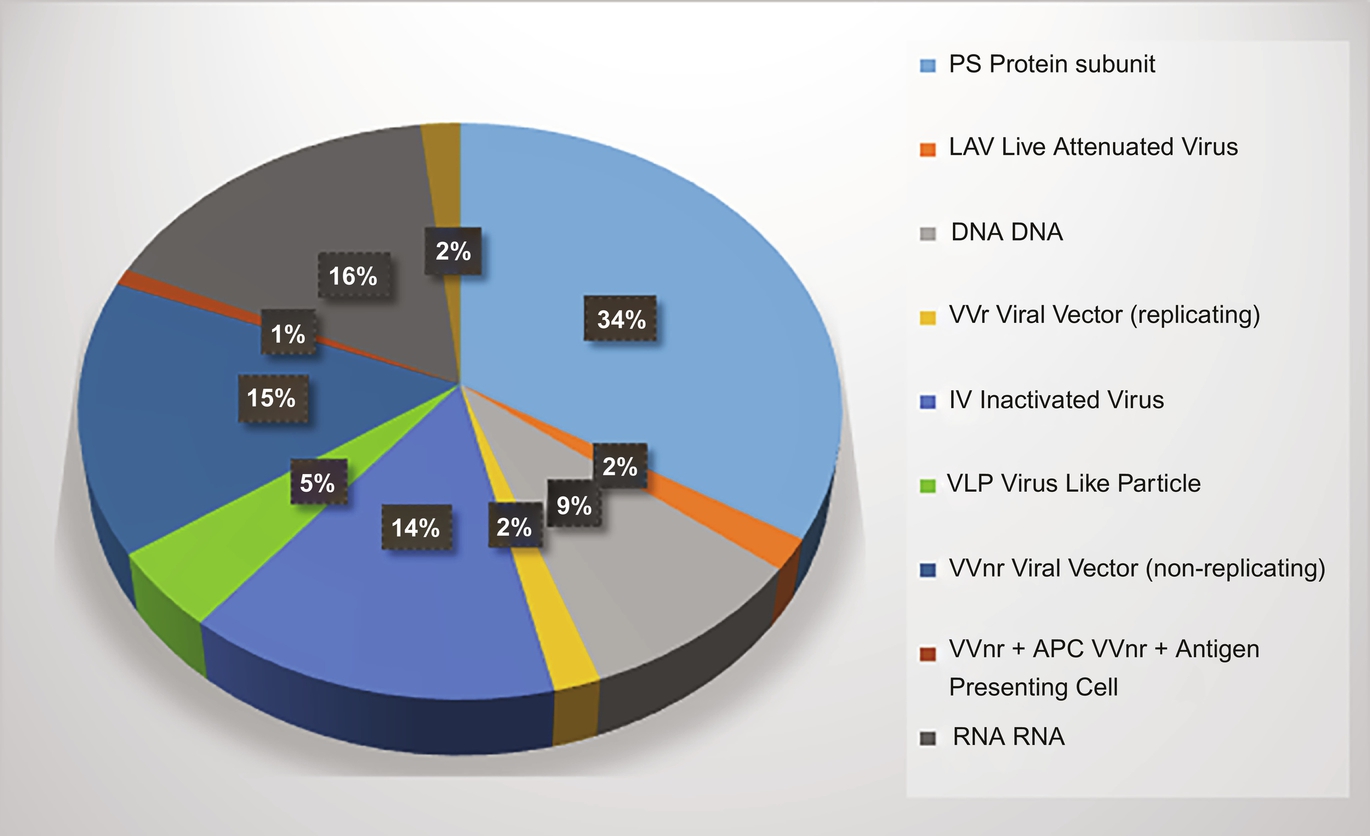 Fig. 4 Percentage representation of vaccine candidates in clinical development. Given is a statistical representation of the different vaccine candidates currently under clinical development.
Fig. 4 Percentage representation of vaccine candidates in clinical development. Given is a statistical representation of the different vaccine candidates currently under clinical development.
4.4: Virus-like particle vaccines
Hepatitis B and human papillomavirus vaccines are based on this platform. Virus-like vaccines are subunit vaccines that are designed to closely resemble the structure of a virus and can induce robust immune responses to the antigen(s) expressed on their surface; they have good safety profiles since they do not contain the pathogen's genetic material. Virus-like particle (VLP) vaccines make use of the immunogenic and safety property of empty virus particles with multiple copies of the same antigen on their surface. The presence of multiple copies of antigen induces a stronger immune response than a single copy. Virus-like particles are formed when proteins S, M, and E of enveloped coronaviruses, with or without N, are co-expressed in eukaryotic cells (Lokugamage et al., 2008). The viral particle detaches from the eukaryotic producer cells through budding. The VLPs produced are identical in structure to the pathogenic virus but lack a viral genome and are thus non-infectious. When VLPs are administered, they bind to ACE2 + cells through their surface S protein in the same manner as the parent virus (Naskalska et al., 2018). The complexity of the production process of VLPs is a challenge in their development. Also, VLPs require an adjuvant and repeated administration just like subunit and inactivated viral vaccines. Nevertheless, the technology used in the production of VLPs is well established, the biology and safety profile of coronavirus VLPs are understood. Five virus-like particle candidates for SARS-CoV-2 produced are currently in clinical trials as shown in Table 1 (WHO, 2020).
4.5: Vectored vaccines
Vectored vaccines are one of the newer platforms for vaccine development. Vectored vaccines are designed using harmless viruses to deliver the pathogen's genetic materials to recipient host cells to produce antigenic proteins to stimulate immune responses. They are modified versions of different viruses with reduced virulence and replication potential but maintain their capacity to infect human cells. Commonly used vectors which are effective in eliciting strong immune response are adenovirus, measles, and vesicular stomatitis virus (VSV) vectors. Recombinant viral-vectored vaccines are developed as either replicating viral vectored vaccines or non-replicating viral vectored vaccines (Jeyanathan et al., 2020). The non-replicating type is incapable of self-propagation but has the advantage of reduced adverse effects. The replicating type, though attenuated, retains its ability to make new viral particles. Hence they can provide vaccine antigen for a longer time. Consequently, a lower dose of the vaccine may be enough to generate a robust immune response. Conversely, non-replicating vectors should be administered in higher dosages since they are incapable of forming new antigens (Jeyanathan et al., 2020). The replication-deficit viral platforms are mostly based on adenovirus or MVA, and most of these vaccine candidates express the S protein or RBD of SARS-CoV-2. Replication- competent viral vectors are mainly based on VSV. The presence of existing antibodies against the viral backbone in people previously exposed to the vector can reduce the magnitude of the triggered immune responses. In line with this, administration of booster doses where it is needed becomes challenging as antibodies against the viral vector produced after the prime vaccination can reduce the immunological response. This problem can be circumvented by using a viral vector with low human sero-prevalence such as ChAd (adenovirus derived from chimpanzees) (Jeyanathan et al., 2020). Although this strategy for vaccine development is new, it has been widely investigated for use in infectious diseases and cancer. The benefits of this platform include safety and the ability to induce strong T-cell responses without the need for an adjuvant (Draper & Heeney, 2010). Replicating viral vector vaccines need to be administered only once for protection and have a natural tropism for the respiratory mucosa meaning that they are suited for respiratory tract vaccination (Afkhami, Yao, & Xing, 2016). There is already existing technology for their large-scale manufacturing and storage. Recombinant viral vectors are one of the most common platforms for COVID-19 vaccine development, with eighteen candidates currently in clinical trials (Fig. 4).
4.6: Nucleic acid vaccines
Nucleic acid vaccines are based on recent trends in vaccine development and involve the delivery of genetic code for in situ production of viral antigen. They provide the genetic instructions to the cells in the body and, in turn, the cells translate the information into proteins which then stimulate an immune response (Jackson, Anderson, et al., 2020; Jackson, Kester, Casimiro, Gurunathan, & DeRosa, 2020; Pardi, Hogan, Porter, & Weissman, 2018). Nucleic acid vaccines are strong inducers of both humoral and cellular adaptive immune responses, and they are easy to use because all you need is a genetic sequence that encodes for a viral antigen and a delivery platform to make them. The potential of mRNA vaccine is seen in previous studies of influenza, rabies, and Zika virus infections in animals (Bahloul, Lassoued, & Sfar, 2014; Chahal et al., 2016; Pardi et al., 2018; Petsch et al., 2012; Schnee et al., 2016). The antigen-encoding mRNA is complexed with a carrier such as lipid nanoparticles. The lipid carrier protects the mRNA when it first enters the body and also helps it to get inside cells by fusing with the cell membrane. This ensures that the nucleic acid is efficiently delivered in vivo into the cytoplasm of host cells where protein translation occurs (Lutz et al., 2017; Pardi et al., 2018). mRNA vaccines are non-infectious and are synthesized by in vitro transcription, free of microbial molecules. These advantages distinguish mRNA vaccines from other vaccines in terms of safety, effectiveness, and anti-vector immunity, allowing for their rapid and low-cost manufacturing as well as repeated vaccination. mRNA-1273, which is a COVID-19 vaccine candidate produced by Moderna, and encodes a prefusion stabilized SARS-CoV-2 S protein encapsulated in lipid nanoparticles. The phase I clinical trial data indicate that low and medium doses of two repeated parenteral injections are generally safe and induce strong S protein-specific antibody responses and a primarily CD4 + T cell response in most trial participants (Jackson, Anderson, et al., 2020; Jackson, Kester, et al., 2020). The clinically advanced Pfizer and BioNTech COVID-19 vaccine is also based on this platform. One of the major drawbacks of mRNA vaccines is the storage temperature of between − 70 °C and − 20 °C which complicates the distribution logistics of these vaccines especially in developing countries. There are eighteen mRNA-based COVID-19 vaccines in clinical trials (Fig. 4). Plasmid DNA vaccines share several characteristics with mRNA vaccines, including safety, ease of production, and scalability (Hobernik & Bros, 2018). DNA molecules are more stable than mRNA and can be stored at 4 °C, thereby simplifying the storage and distribution of this type of vaccine. Some of the major limitations of DNA vaccines are poor immunogenicity, repeated administration, and the need for an adjuvant. Currently, there are ten DNA-based COVID-19 vaccines in clinical trials as shown in Fig. 4, with some examples represented in Table 1.
5: Relevant SARS-CoV-2 antigen explored in the design of vaccines
5.1: Structural, sub-structural, and non-structural proteins
SARS-CoV-2, like other CoVs, has four major structural proteins, including the spike (S) protein, the envelope (E) protein, the membrane (M) protein, and nucleocapsid (N) protein (Shin et al., 2020; Wu et al., 2020). Apart from these four structural proteins, the CoV viruses also encode specific and sub-structural proteins like hemagglutinin-esterase (HE) protein, 3a/b protein, and 4a/b protein (Zarandi, Zinatizadeh, Zinatizadeh, Yousefi, & Rezaei, 2021). CoVs genomes also have non-structural proteins which are more protected than structural proteins. Additionally, compared to non-structural proteins, structural proteins easily undergo mutation to suit adaptation to the host's system (Zarandi et al., 2021). Apart from their role in replication, the other functions of non-structural proteins are yet to be understood (Chen, Liu, & Guo, 2020).
5.2: Spike (S) protein
The diverse roles played by the structural proteins, especially S protein, make them target sites for vaccine development. The S protein is the most researched of all structural proteins because it plays a vital role in the entry of SARS-CoV-2 into host cells at the infection stage.
Just like other CoV viruses, the SARS-CoV-2 surface possesses homotrimers of S protein which are fashioned as spikes used for binding to host receptor cells (Nieto-Torres et al., 2014; Zarandi et al., 2021). The S protein is made up of two sub-units: S1 and S2 (Fig. 5). While S1 controls binding to ACE2 on the host cell, S2 is responsible for the fusion of the host cell membrane during the process of infection (Xia et al., 2020). The receptor-binding domain (RBD) is located in S1, and this is where the infection begins. Endocytosis of SARS-CoV-2 occurs with the binding of RBD to ACE2 receptor (Fig. 6), followed by the release of the viral pack in the host cell cytoplasm after the interaction with endosomal proteases. In fact, research has pinned the high transmission rate of SARS-CoV-2 to the high affinity of its RBD to bind to ACE2 receptors, an affinity that is 10–20 times higher than that of the SARS-CoV virus. Cytokine storm which is a characteristic severe immune response in COVID-19 disease is as a result of the endocytosis of SARS-CoV-2 into lung cells (Zarandi et al., 2021).
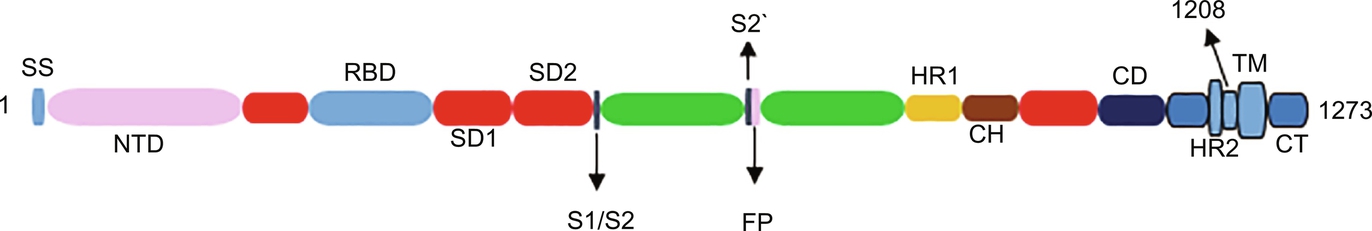 Fig. 5 Spike protein structure in SARS-CoV-2. SS, signal sequence; S2′, protease cleavage site; FP, fusion peptide; HR1, heptad repeat 1; CH, central helix; CD, connector domain; HR2, heptad repeat 2; TM, transmembrane domain; CT, cytoplasmic tail. Adapted from Kheirandish et al., 2021.
Fig. 5 Spike protein structure in SARS-CoV-2. SS, signal sequence; S2′, protease cleavage site; FP, fusion peptide; HR1, heptad repeat 1; CH, central helix; CD, connector domain; HR2, heptad repeat 2; TM, transmembrane domain; CT, cytoplasmic tail. Adapted from Kheirandish et al., 2021.
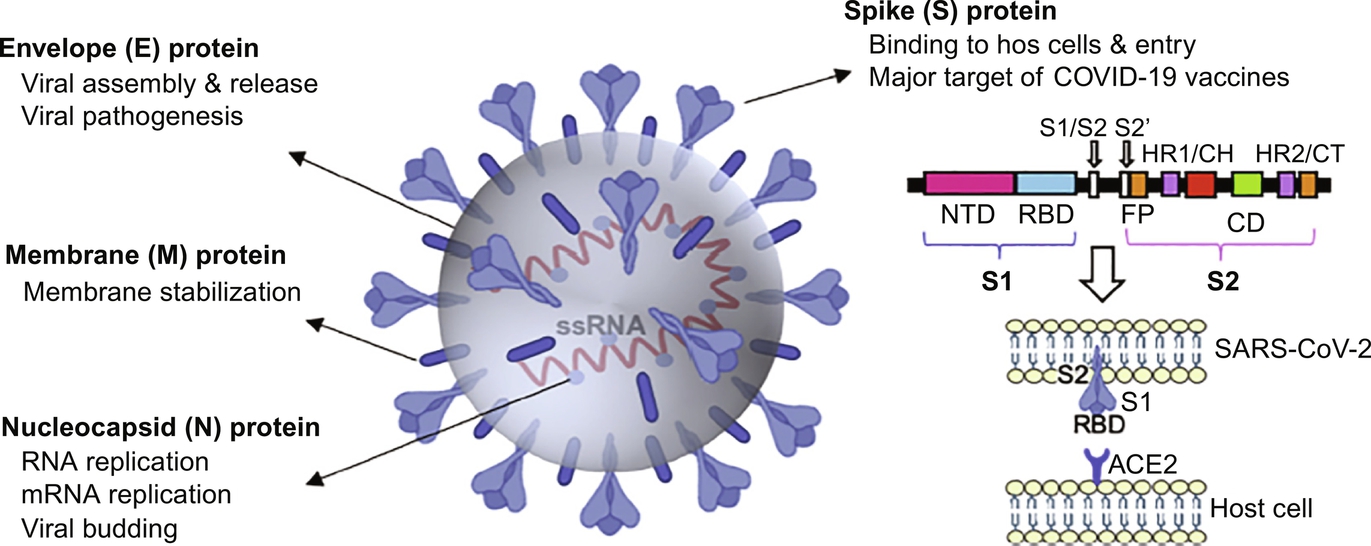 Fig. 6 Diagrammatic representation of the SARS-CoV-2 structural proteins (Young et al., 2021). Reproduced by permission of the Elsevier Permissions.
Fig. 6 Diagrammatic representation of the SARS-CoV-2 structural proteins (Young et al., 2021). Reproduced by permission of the Elsevier Permissions.
Currently, most pre-clinical and clinical researches on vaccine development are based on S protein. About 35 of the 47 vaccine candidates undergoing clinical trials are based on S protein. Components of the S protein exploited as antigens include the full-length S protein, the RBD domain, the S1, and S2 subunits, the N-terminal domain (NTD), and the membrane fusion peptide (Ahmad et al., 2021). The information acquired during SARS and MERS vaccine development in 2002 and 2012, respectively, as well as the knowledge about the structure of SARS-CoV-2, has revealed that S1, S2, and RBD possess epitopes that can be targeted for the induction of neutralizing antibodies. Analysis of the sera of a cohort of convalescent SARS-CoV-2 patients revealed the presence of neutralizing antibodies (NAbs) acting on the S1, RBD, and S2 sites of the virus (Wu et al., 2020). A good number of these patients (95–100%) demonstrate neutralizing activity 14 days after the onset of COVID-19 symptoms. The neutralizing activities exhibited include IgM, IgG, and antibodies targeting RBD of SARS-CoV-2 (Suganya, Divya, & Parani, 2021; Zhao et al., 2020). NAbs elicit humoral responses in the host immune system, inhibiting the SARS-CoV-2 binding to the human receptor, in addition to instigating viral lysis through antibody-mediated opsonization or complement activation (Jiang, Hillyer, & Du, 2020; Suganya et al., 2021).
Another approach proposed for this target site is the development of glycosylated SARS-CoV-2 S vaccines since computational studies have revealed that glycosylated SARS-CoV-2 S proteins have more organized conformation compared to non-glycosylated forms of the protein (Banerjee, Santra, & Maiti, 2020). This may be because the SARS-CoV-2 S protein is extensively glycosylated in situ (Shin et al., 2020). Researchers are also exploiting the receptor binding affinity of the RBD domain of the S1 subunit of the S protein. Studies have revealed that SARS-CoV-RBD can generate antibodies that cross-react with S2-RBD protein, leading to SARS-CoV RBD-actuated antisera production. The SARS-CoV-RBD actuated antisera cross-kill SARS-CoV-2. Hence, the investigation into possible production of SARS-CoV RBD-based vaccine are being advocated (Ahmad et al., 2021; Tai et al., 2020). ChADOx1 vaccine candidate containing optimized full-length surface S glycoprotein with a tissue plasminogen activator is the most advanced S protein-based vaccine candidate. In ChADOx1, a replication-deficient chimpanzee adenovirus was used as a carrier to deliver a SARS-CoV-2 S protein to elicit a protective immune response. A single dose of ChADOx1 in 6 rhesus macaques prevented the development of SARS-CoV-2 induced pneumonia (Ahmad et al., 2021; van Doremalen et al., 2020).
Nanotechnology has been employed in the design of S protein-based vaccine candidates. The Moderna and Pfizer/BioNTech vaccines have lipid nanoparticles as carriers. The mRNA which codes the S protein of the SARS-CoV-2 (Polack et al., 2020) is loaded into phospholipid membranes (De Soto, 2021). The introduction of the lipid-based nanoparticles into the host system results in the fusion of its phospholipid membrane with the host membrane and the discharge of the mRNA into the cytoplasm of the target cell. Translation of the mRNA of the S protein occurs at the rough endoplasmic reticulum, followed by the production of the S protein within the cytoplasm. Degradation and expression of the S protein by Major Histocompatibility Complex I (MHC I) and II (MHC II) then occurs. MHC II is usually present in antigen-presenting immune cells such as macrophages, B-cells, and dendritic cells. The S protein fragment is then presented by the MHC II molecule to a T-helper cell, after which the S protein binds to the T-helper cells with its T-cell receptor protein, as well as binds the MHC II molecule using its CD4+ receptor (Shin et al., 2020). These reactions cause T-helper cells to release interleukins which causes B cells to proliferate. In addition to the proliferation of B cells, is its differentiation into plasma cells, which then releases specific antibodies to the S protein fragment. The interleukins, in turn, cause the initially produced T-helper cells to proliferate and form T-helper memory cells. Both the T-helper cells and plasma cells produce antibodies against some domains of the S protein of SARS-CoV-2 during future attacks (De Soto, 2021). Furthermore, through the MHC I complex, cytotoxic T cells are able to bind S protein fragments expressed by non-immune cells. This interaction leads to the release of cytokines by cytotoxic cells and the proliferation and differentiation of B cells into plasma cells. Additionally, the interaction of the S protein with cytotoxic T cells paves the way for the elimination of cells infected by the S protein of the SARS-CoV-2 virus in the future (De Soto, 2021).
5.3: Membrane (M) protein
Unlike the S protein, few studies have focused on SARS-CoV-2 M protein (S2M) (Suganya et al., 2021). The function of the M protein starts with the assembly of viral particles and the initiation of its budding process (Ahmad et al., 2021; Zarandi et al., 2021), as well as membrane stabilization functions (Young et al., 2021). M protein is located amid the S and E proteins in the virus envelope. During the process of viral particle formation, S2M interacts with N, E, and S proteins as well as with its self (Ahmad et al., 2021). Recent studies in SARS-CoV-2 recovered patients have revealed the existence of appreciable CD4+ and CD8+ T cells initiated immune responses against SARS-CoV-2 M protein (Grifoni et al., 2020). Studies carried out in the past on immunized rabbits have also revealed that synthetic peptides obtained from immunodominant epitopes prompted significant antibody-induced immune reactions, and further highlighting the immunogenicity of S2M (He et al., 2004). OncoGen (a SARS-CoV-2 vaccine company) has designed a synthetic long peptide vaccine candidate to target the spike and membrane proteins of the SARS-CoV-2 virus. However, the results of the pre-clinical studies are still being awaited (Ahmad et al., 2021).
5.4: Nucleocapsid (N) protein
N protein functions in the area of SARS-CoV-2 replication (Fig. 6). During viral assembly, N protein which is located within the viral envelop encloses the RNA to shape the helical nucleocapsid. Nucleocapsid protein can connect with the RNA virus genome through its two domains, but through different mechanisms (Zarandi et al., 2021). None of the SARS-CoV-2 vaccine candidates in clinical trials is based on N protein alone. However, a Human Adenovirus Type 5 Vector (hAd5) in Phase I clinical trial, and developed by ImmunityBio, Inc. & NantKwest Inc. encodes S and N proteins (WHO, 2020). On the other hand, a vaccine candidate designed to target the spike S1 subunit, membrane protein, and N protein is being developed by the National Research Centre in Egypt, however, it is still undergoing pre-clinical trials (Zarandi et al., 2021).
5.5: Envelop (E) protein
The E protein is involved in the pathogenesis of the SARS-CoV-2 virus disease through its role in the release of the virus and ion channel activity (Karpiński, Ożarowski, Mrozikiewicz, & Wolski, 2021; Zarandi et al., 2021). Unlike other structural proteins, the E protein is not a usual target for vaccines since they are not sufficiently immunogenic because of its structure and low quantity (Karpiński et al., 2021; Young et al., 2021). Nonetheless, a recent study by Abdelmageed and co-researchers targeted a multiepitope-based peptide vaccine against the E protein of human COVID-19 (Abdelmageed et al., 2020). There is a need, however, to thoroughly investigate the vaccine to determine its immunogenicity and safety profile (Ahmad et al., 2021).
6: Nano-based strategies for COVID-19 vaccine development
Various nano-sized formulations have been explored in the development of vaccines over the years. The use of nano-based systems in the development of vaccines has shown several benefits over conventional molecular formulations. These benefits include the ability of the nano-systems to deliver various types of vaccines while ensuring the protection of the encapsulated antigens from premature degradation and non-target sites like the macrophages. (Alimardani, Abolmaali, & Tamaddon, 2021). This is largely because the systems are designed in a manner that permits the proper exposure of the antigen at the same time protecting them from proteases and nucleases in the body (Ruiz-Hitzky et al., 2020). Also, the fact that nanotechnology permits the design and development of nano-sized materials with the required physicochemical and biologic properties such as size, morphology, solubility among others, makes it highly useful in the development of vaccine delivery systems. Surface modification of these nano-sized materials with targeting moieties has been known to increase uptake by phagocytic APCs (Alimardani et al., 2021). These systems have also been successfully engineered to reduce antigen toxicity while improving the immunogenicity of antigens and adjuvants. They can also act as adjuvants themselves potentiating the immune responses to vaccines (Alimardani et al., 2021). Some of these systems also have the advantage of antigen delivery via alternative routes than the parenteral routes such as the oral, nasal as well as transdermal routes (Alimardani et al., 2021). These nano-sized formulations have also been employed to ensure thermo-stability, thus improving vaccine distribution and decreasing vaccine failures (Alphandéry, 2020). In addition to thermo-stability, the nano-size confers on these preparations the ability for easy targeted uptake in desired cells or tissues thus triggering an optimal immune response against the virus (Shin et al., 2020).
6.1: Nano-carriers for antigen delivery
The use of nano-carriers for antigen delivery is usually done in two ways; by either loading the antigens inside or on the surface of the nanocarriers. The choice between the two options usually depends on the antigens' biological stability, physicochemical characteristics, target sites, and required immunogen release rate (Alimardani et al., 2021).
6.1.1: Polymeric nano-delivery systems
Polymers have been explored extensively in the world of drug design and delivery with wide applications. Biocompatibility, as well as biodegradability, are two key features highly sought after in the selection of polymers for drug delivery. This supports the extensive use of poly (D, L-lactic-coglycolic acid PGLA) and poly (D, l-lactide-co-glycolide PLG) in the design and development of nano polymeric carriers for drug delivery. (Zhao et al., 2014). These systems have also been employed as delivery systems for vaccines because of their ability to ensure the controlled release of antigens and adjuvants (Kim et al., 2014). In addition, the surface and particle size of these particles have been modified in vaccine development to encourage oral, mucosal as well as systemic delivery of antigens (Kim et al., 2014). Nano-polymeric systems composed of natural polymers such as chitosan have been studied in mucosal vaccine delivery systems. Chitosan nanoparticles encapsulating hemagglutinin-split influenza were prepared by ionic cross-linking of the chitosan polymer in the presence of sodium tripolyphosphate. On administration of the two doses via the nasal route, the chitosan nanosystems were able to produce antibody responses compared to the response generated by the hemagglutinin-split influenza virus alone (Kim et al., 2014).
6.1.2: Lipid-based nano-delivery systems
Lipid-based nano-systems such as liposomes have extensively been explored in the design and development of therapeutics as well as vaccines. They are one of the most promising approaches in nanotechnology designed for encapsulating as well as the targeted release of antigens in vaccine development (Raoufi, Bahramimeimandi, Salehi-Shadkami, Chaosri, & Mozafari, 2021). Their use in developing vaccines received an increased level of attention after their immunomodulatory actions were discovered. Since then, many other reports have shown that their physicochemical characteristics such as size, structure, and lipid components can be modified to enhance immunogenicity (Kim et al., 2014). These systems are very efficient in encapsulating DNA or RNA-based immunogens. About 10 COVID-19 vaccine candidates make use of lipid-based nano-systems as carriers (Malabadi, Meti, & Chalannavar, 2021).
Liposomes are made up of biocompatible phospholipid bilayers and this confers on them the ability to deliver hydrophobic as well as hydrophilic moieties. This property is particularly important in vaccine delivery as it could permit the simultaneous delivery of antigens and adjuvants. In addition to this, the surface of liposomes can be modified using appropriate functional moieties (such as lipid components of the lipid bilayer) and targeted to immune cells, thus enhancing immune responses and consequently efficacy of the vaccine (Kim et al., 2014). Reports have shown a combination of dimethyl dioctadecyl ammonium (DDA) liposomes to enhance immunity against influenza, chlamydia, and tuberculosis infections (Alving, Beck, Matyas, & Rao, 2016; Christensen, Smith, Andersen, & Agger, 2011). The mRNA-1273 vaccine developed by Moderna is a liposomal formulation encapsulated with nucleoside modified mRNA that encodes the SAR-COV-2 S glycoprotein. The mRNA-1273 induced anti-SARS-COV-2 responses in all participants without significant toxicity (Petkar et al., 2021). Also, reports from the pre-clinical trials showed a 94.1% efficacy in preventing COVID-19 after administration of the two-dose regimen (Petkar et al., 2021). Also, the BNT162b2, by Pfizer and BioNTech is basically a lipid nanoparticle encapsulating nucleoside modified RNA vaccines with encoded membrane-anchored SARS-COV-2 S protein. It was reported that the two-dose vaccine regimen was 95% effective against COVID-19 infections, with mild to moderate side effects. NVX-CoV2373 is another nanoparticulate vaccine candidate, made of trimeric full-length recombinant S-protein of the SARS-COV-2 and MATRIX-M1 adjuvant. Studies conducted so far showed it was able to elicit anti-spike IgG antibodies which possessed ACE2 receptor blocking as well as virus neutralization capabilities (Magnusson et al., 2018; Rao et al., 2020). In addition to this, NVX-CoV2373 also induced T and B cell responses and further clinical evaluation produced results that support the safety of this vaccine candidate (Pandey et al., 2021).
A study by Zhang et al. explored the use of ionizable lipid-based nanoparticles for the delivery of mRNA encoding the receptor-binding domain (RBD) of SARSCoV2. The system was made up of ionizable lipid (1,2-distearoyl-sn-glycero-3-phosphocholine), cholesterol, and pegylated lipids combined in various ratios. The lipids were dispersed in ethanol and combined with the mRNA contained in a citrate buffer of pH 4.0, using a T-shaped microfluidic mixer. The nanoparticles were formed through nano-precipitation (Zhang et al., 2020). On administration of this vaccine protocol by IM, the authors observed that there was protein expression at the site of the administration as well as induction of significant SARS COV2 specific IgG and neutralizing antibodies. The formulation was able to induce SARS COV2 specific CD4+ and CD8+ effector memory T cells in the spleen. In addition, on storage, the lipid nano-formulation was stable at room temperature for up to 7 days (Zhang et al., 2020). Another study by McKay et al. (2020) developed a vaccine strategy of lipid nanoparticles encapsulating SARSCOV2 spike protein-encoding self-amplifying RNA (saRNA-LNP). This formulation was observed to induce significant SARSCOV2 specific IgG in mice as well as higher cellular response and viral neutralization (McKay et al., 2020). These are pointers that this could be a very robust vaccine candidate for the prevention of COVID-19.
6.1.3: Inorganic nano-delivery systems
In addition to their contributions in diagnostics, several inorganic nanoparticles, have recently been considerably explored in the area of vaccine development and delivery. This is mainly because of their rigid structure as well as ease of synthesis. (Zhao et al., 2014). In addition, the fact that these nano-systems possess smaller particle sizes, have improved stability and permeability as well as high drug loading capacity have made them ideal for drug delivery. Also, inorganic nanoparticles have been known to exhibit high cellular uptake, non-immunologic responses, low toxicity, and distinct physicochemical and biological properties compared to their bulk counterparts. These properties make them ideal for antigen delivery in vaccine development (Poon et al., 2018). Recently, hybrid inorganic nano-systems have been developed which comprises an inorganic core and an organic outer shell to improve their safety in vivo (Poon et al., 2018). Gold nanoparticles of various shapes, sizes, and surface modifications have been widely used as carriers for antigens from viruses such as the influenza virus (Zhao et al., 2014). Silica-based nanoparticles possess immense potentials in the area of vaccine design, development, and delivery. This is primarily due to the fact that they are remarkably biocompatible. Also, the presence of abundant surface silanol groups, on modification, makes for the introduction of additional functionality like improved cellular recognition and uptake, better interaction with cells as well as improved absorption of specific biomolecules. These systems have also been reported to induce very strong humoral in addition to cell-mediated responses, features that have greatly influenced their use in vaccine development (Ghaffari et al., 2021). Nuvec®, a silica nanoparticle (combined with polyethyleneimine), developed by N4 Pharma, has been reported to protect and aid the delivery of RNA/DNA antigens to targeted cells. In addition, Nuvec® also possesses high loading capacity, strong bonding, and high cellular uptake. A study by Theobald (2020) strongly suggested that Nuvec® could be employed as a non-viral delivery vehicle for vaccines and hence could be a safe and effective alternative to lipid nanoparticles and non-replicating viral vectors in the development of the SARS-COV2 vaccine. Erasmus et al. (2020) developed an alphavirus –derived replicon RNA vaccine candidate (repRNA-COV2S), comprising of SARS-COV-2 S-protein replicons in a squalene-based emulsion of lipid inorganic nanoparticles (LION). The inorganic nanoparticles were composed of an inorganic (Superparamagnetic iron oxide, Fe3O4), and a cationic lipid component (1,2-dioleoyl-3-trimethylammonium propane). Animal studies conducted in mice and macagues revealed that the vaccine was able to produce antigen-specific antibody responses, which were comparable to the convalescent response from COVID-19 (Erasmus et al., 2020). Another remarkable property of this vaccine candidate was its ability to elicit robust antibody responses, producing neutralizing antibodies in young and old mice alike, with the administration of just one dose (Erasmus et al., 2020). In addition, the strategy of manufacturing the vaccine in two vials-one of each containing the LION and the other the replicon RNA components, makes the scale up of the vaccine formulation easier as incompatibilities between the components is avoided (Erasmus et al., 2020).
6.1.4: Carbon-based nanomaterials
Carbon-based nanomaterials (CBNs) are fast becoming highly relevant material due to the existence of diverse allotropes of carbon, from renowned allotropic phases such as amorphous carbon, graphite, and diamonds to newly discovered auspicious carbon nanotubes (CNTs), quantum dots (QDs), and fullerene (Zhang et al., 2017). As each member of the carbon family exhibits inimitable features, they have been widely used in diverse biological applications including biosensing, drug delivery, tissue engineering, imaging, diagnosis, and cancer therapy (Bhattacharya et al., 2016). Carbon nanotubes (CNTs) classified into single-walled carbon nanotube (SWCNT) and multi-walled carbon nanotube (MWCNT) are hollow cylinders consisting of graphitic sheets. These systems are recently receiving a lot of attention in the area of vaccine development and delivery (Kim et al., 2014). This is because these systems are capable of carrying multiple antigens, are easily taken up by antigen-presenting cells, and display low toxicity. However, they are largely insoluble and non-degradable, hence it is vital to determine how this will affect the safety of the carbon-based nano delivery systems. Several reports have also illustrated the possibility of using carbon-based nanosystems for the oral delivery of antigens (Kim et al., 2014).
6.2: Nano-vaccine adjuvants
The development of vaccines has been one of the most successful tools in the control of several infectious diseases. However, studies have shown that some of these vaccines generate rather weak immune responses. This is as a result of the fact that these vaccine antigens are made with only certain parts of the pathogens; hence, the need for adjuvants to aid the provocation of robust immune responses (Petkar et al., 2021). The inclusion of an adjuvant in a vaccine formulation has been observed to improve the immunogenicity of antigens while decreasing the concentration of antigens as well as the number of doses required to induce protective immunity. These substances have also been known to improve the efficacy of vaccines in newborns, the elderly as well as people with compromised immune systems (Petkar et al., 2021).
Nano-emulsions have been widely developed over the years as adjuvants (Zhao et al., 2014). These systems are composed of either oil-in-water or water-in-oil emulsions, either carrying the vaccine antigens in their core or simply mixed with them (Zhao et al., 2014). The MF59® is a type of oil-in-water emulsion used as a potent and safe adjuvant in vaccine design and development. The adjuvant effect of the MF59® has been widely evaluated in the development of influenza vaccines among others (De Donato et al., 1999; Hagan, 2007). The Matrix-M®, developed by Novavax Inc., is a nano-based saponin-derived adjvant, comprising of saponins from the tree Quillaja saponaria, mixed with cholesterol and phospholipids. These substances combine to form stable nanoparticles that can easily be incorporated into various formulations of vaccine antigens. Studies have shown that the inclusion of this adjuvant can precipitate an antigen dose reduction thereby resulting in the reduction of cost of production, ultimately leading to the production of a cost-effective vaccine (Machhi et al., 2021). Recently, the Matrix-M was employed as an adjuvant in a COVID-19 vaccine candidate, NVX-CoV237, also developed by Novax Inc. The Matrix –M improved the recruitment of APCs at the injection site, thus causing an increase in T-cell activation in the lymph nodes (Keech et al., 2020).
CoVaccine HT®, an oil-in-water nano-emulsion made of negatively charged sucrose fatty acid sulphate ester and squalene, was evaluated as an adjuvant with the SARS-CoV-2 S-protein in mice (Bonam et al., 2021). This research was fuelled by the fact that the CoVaccine HT® was successfully used in different vaccine formulations including malaria, ebola, zika among many others (Bonam et al., 2021; Kusi et al., 2011; Lehrer et al., 2018). The study revealed that CoVaccine HT® induced significantly higher antigen-specific antibody titers, cell-mediated immune responses, as well as virus-neutralizing antibodies, compared to alum, which is the gold standard of conventional adjuvants (Kusi et al., 2011). In addition to nano-emulsions, other nano-based adjuvants have also been developed for application in the development of COVID-19 vaccines. A recent study by Rao et al. (2020) designed and developed nano decoys possessing cellular membrane nanovesicles derived from genetically modified 293T/ACE2 and THP-1 cells. These nano-decoys possess bio-properties such as abundant ACE2 and cytokine receptors, which basically compete with the host cells, thereby interfering with the process of viral infection and replication. Adavax, a microcrystalline polysaccharide adjuvant derived from delta inulin, has also been included in a COVD-19 vaccine candidate developed by a molecular modelling approach, COVAX-19. The adjuvant was observed to improve the immunogenicity of the vaccine antigen (Rao et al., 2020).
6.3: Delivery devices
The development of microneedles has become a very powerful tool in the world of vaccine design and development. This is primarily because these devices have the ability to elicit a sustained release of vaccine antigens, thus eliminating the need for multiple doses. This is the case especially with microneedles made with sustained-release polymers or embedded with nanoparticles. (Menon et al., 2021). They can also be self-administered, thus eliminating the need for a trained health professional. In addition to these benefits, microneedles have been reported to induce robust immune responses against different pathogens. This is due to the presence of abundant dermal dendritic cells among other immune systems in the skin (Menon et al., 2021). In a pandemic, such as the one caused by COVID-19, where mass immunization program is one of the key tools in the control of the spread of the SARS-CoV2, the development of microneedles shows a lot of promise in the actualization of mass immunization. Several companies have developed or repurposed their microneedles for use in the control of the COVID-19 pandemic. The use of microneedle patches for the administration of COVID-19 vaccines is also being proposed to solve the problem of cold chain storage. The presence of a good number of antigen-presenting cells on the epidermis and dermis layer of the skin, which is likely to reduce the dose of vaccines, is included as one of its advantages. There are chances that microneedle-based formulations may be cost-effective since they may be able to reduce the need for cold chain storage (O'Shea, Prausnitz, & Rouphael, 2021; Tran et al., 2020). A study carried out by Kim et al. succeeded in formulating a dissolvable microneedle matrix delivering SARS-CoV-2 antigen, and it was tested in a mice model. Vaccine-induced antibodies were generated in significant amounts within 2 weeks post-administration (Kim et al., 2020). In another study, the RBD domain peptide of the SARS-CoV-2 spike protein was delivered through a microneedle-based formulation to mice. An ample amount of T-cell response, as well as antibody generation, was also reported (Kuwentrai et al., 2021). More research is still needed to determine procedures necessary for upscaling production and commercialization of a microneedle-based formulation as this will be very beneficial in developing countries of the world (Kumar & Kumar, 2021).
6.4: Novel alternative routes of administration
The role of nanotechnology in the area of vaccine development as established earlier is highly invaluable. In addition to the benefits of providing suitable carrier systems for antigen, acting as adjuvants of stellar quality as well as having a huge impact on the development of novel delivery devices for a vaccine, nanotechnology has provided the opportunity for the exploration of various routes for vaccine delivery by precipitating the birth of a plethora of formulations with various remarkable characteristics. The determination of the most suited route of administration is a very vital aspect of vaccine design and development. This is particularly important as vaccine formulations, more than other pharmaceutical formulations, are hugely affected by the route of administration, as this can determine the extent and quality of immune responses (Wang, Hu, et al., 2020; Wang, Peng, et al., 2020; Wang, Zhao, et al., 2020). Certain reports have suggested that establishing mucosal immune protection by mucosal vaccination either via pulmonary, oral, or intranasal routes, might be more preferable for preventing COVID-19 since the disease is predominantly a respiratory disease-causing immense pulmonary inflammation (Wang, Hu, et al., 2020; Wang, Peng, et al., 2020; Wang, Zhao, et al., 2020). Formulations administered via these routes come in direct contact with the affected epithelial cells in the lungs, thereby bringing about a fast onset of action which could, in turn, decrease the symptoms of respiratory distress as well as lung occlusions (Abdellatif, Tawfeek, Abdelfattah, El-Saber Batiha, & Hetta, 2021). However, the majority of the COVID-19 vaccine candidates are administered via the parenteral routes, primarily the intramuscular route in order to achieve prolonged release of the antigen and bring about higher chances of interaction and uptake by an antigen-presenting cell (Young et al., 2021). That notwithstanding, when vaccines are administered through this route, it confers potentially only non-mucosal systemic immunity and thus leaving questions as regards the durability and efficacy of the mucosal immunity after vaccination, which is of great importance in the prevention of viral entry through the oro-respiratory tract (Ashraf et al., 2021). In addition, the nasal and pulmonary routes are also being projected for the administration of SARS-CoV-2 vaccines because studies have revealed that resident memory T cells are detected in lung tissues and airways after recovery from respiratory viral infections (Hogan et al., 2001) which may produce longer-lasting immunity against SARS-CoV-2 (Young et al., 2021). Despite this reality, several researchers are continuously working to develop mucosal as well as other vaccines that have alternative delivery routes to conventional ones. The mucosal route of administration offers the benefit of a lower risk of systemic adverse effects of the vaccine formulation and a needle-free vaccination, making them more patient-friendly and also eliminating the need for a skilled health professional (Strizova, Smetanova, Bartunkova, & Milota, 2021). Some reports have postulated that since the nasal cavity is a major entry point for the SARS_CoV 2, the nasal associated lymphoid tissues (NALT), could present a promising target for COVID-19 vaccine delivery (Pandey et al., 2021). The NALT, majorly consisting of dendritic cells, macrophages, and lymphoid follicles on the activation have been largely implicated in the clearance of viral pathogens from the mucous layer (Pandey et al., 2021). A study exploring an intranasal vaccine candidate for SARS-CoV reported the induction of Trm cells in the lungs, which are vital for protection from viral infection (Baric et al., 2016). Another study by An et al. (2020) showed that a single dose of adenovirus type 5 vectored vaccine encoding the receptor-binding domain of SARS-COV 2 S-protein administered nasally was able to induce both systemic and local immune responses against the SARS-CoV2 in mice. This was attributed primarily to the induction of mucosal IgA and serum neutralizing antibodies (An et al., 2020). Hassan et al. (2020) also developed a chimpanzee adenovirus-vectored vaccine which encodes a perfusing stabilized S-protein (Chad-SAR-CoV-2-S), for delivery via the nasal route. This formulation was reported to induce very high levels of neutralizing antibodies while enhancing systemic and mucosal immunoglobulin A (IgA) and T-cell responses, thus preventing infection of SARS-CoV-2 in the upper and lower respiratory tracts (Hassan et al., 2020). China is currently testing a COVID-19 vaccine designed as an intranasal spray. This vaccine is composed of weakened flu viruses (H1N1, H3N2, and B) combined with segments of the SARSCoV2 s protein, which mimics infection of respiratory viruses and can stimulate immune responses. This candidate promises to provide the advantage of easy scale-up of industrial production and distribution (Abdellatif et al., 2021).
Also, the oral route of drug delivery being the most preferred due to patient compliance and ease of administration has been studied in the delivery of COVID-19 vaccines. An oral COVID-19 vaccine candidate (VXA-CoV2-1), developed by VAXART, USA, is designed as an orally administered recombinant coated tablet. It comprises an enteric-coated tablet containing an adenoviral vector, encoding for the genes for the S and N-proteins of the SARS-COV2. On administration, VXA-CoV2-1 triggers a mucosal immune response for defence against viruses causing respiratory infections as well high titres of neutralizing antibodies against the SARS-CoV2. Also, the study revealed that there was no sign of mucosal damage or weight loss in the hamsters used. Due to the fact that this vaccine candidate targets the N-protein in addition to the S-protein, it promises to provide better protection against the new viral variants. This has recently received approval for phase II clinical trials (Ashraf et al., 2021). A UK-based company, IsoBio, is currently developing the Oral Pro-COVID-19®. This candidate is a non-replicating viral vector vaccine that expresses the S-protein, designed as a thermally stable capsulated form. This is particularly important as it provides the advantage of self-administration and needle-free application. In addition, the need for cold-chain storage is eliminated as the vaccine is designed as a thermally stable capsule. This makes this candidate especially interesting for developing countries with erratic power supply (Ashraf et al., 2021).
7: Benefits and challenges of nanotechnology in COVID-19 vaccine development
Nanotechnological approaches provide the most effective tools for the control of infectious diseases such as COVID-19, from developing stellar preventive measures, therapeutics, and diagnostics to the design and development of safe and effective vaccination protocols. Scientists all over the world have in the last year battled with the development of suitable vaccines in a bid to curb the spread of the SARSCoV 2 virus and nanotechnology has played a crucial role in providing suitable nano-carriers for antigens, adjuvants, and formulations suitable for delivery via alternative routes (Abdellatif et al., 2021). The use of nano-based delivery systems for the delivery of antigens has paved new pathways and provided several advantages such as antigen stability, sustained/controlled delivery of antigens as well as evasion of immune responses (Malabadi et al., 2021). These systems also provide the advantage of delivering antigens and adjuvants simultaneously, thus resulting in enhanced immunogenicity (Lutz et al., 2017; Malabadi et al., 2021). Also, surface modification of nano-carriers has been employed in designing nano-systems for the targeted delivery of antigens to specific cells to improve immune responses (Kim et al., 2014). In addition, the development of multifunctional nano vaccines could enhance the immune response by ensuring target specificity, effectiveness, and stable delivery of vaccine antigens (Kim et al., 2014). One of the major challenges in the development of COVID-19 vaccines is developing vaccine approaches that stimulate both the T and B cell immunity against the SARS-CoV 2, as well as developing vaccines that will be well suited for all populations especially those with compromised immune systems (Alimardani et al., 2021). Nano-based strategies have been explored in tackling these problems by encouraging the design and development of vaccine systems that induce the immune system optimally. Another major challenge encountered in the development of COVID-19 vaccines is the issue of the cold chain storage requirement of some vaccine preparations. It has been estimated that the cost of cold chain storage alone accounts for about 80% of vaccination costs (Wang, Hu, et al., 2020; Wang, Peng, et al., 2020; Wang, Zhao, et al., 2020). This is highly problematic especially for developing countries where the supply of electricity is highly variable. It is because of this that relevant adjustments to the design of formulation using nanotechnology can address this problem by striving to make vaccine formulations stable at room temperature. A vaccine comprising lipid nanoparticles as delivery vehicles have been modified to stabilize the vaccine antigens being delivered (Wang, Hu, et al., 2020; Wang, Peng, et al., 2020; Wang, Zhao, et al., 2020).
Despite the benefits that nanotechnology offers in vaccine development, there are also some issues of concern. One of the major problems of nano-based systems is the issue of toxicity. There are several molecular mechanisms of toxicity elicited by nanosystems. Some of these systems have been known to interact with cellular DNA, interrupting important enzyme functions and thus causing harm to the organism. There have also been some reports of these systems generating reactive oxygen species that eventually cause harm to genetic materials or disrupt vital enzyme functions (Pandey et al., 2021). Also, because the process of vaccine development testing and regulation is long, taking about 10–15 years, and the current COVID-19 vaccine candidates have been developed within a space of a year, there are concerns about the safety and long term effects of these formulations (Soleimanpour & Yaghoubi, 2021). The use of a multi-component nano-based vaccine, having complex structural make-up (Bonam et al., 2021) can bring about an increased cost of production as a result of a rigorous process of production. Some reports have also argued that since many nano-vaccines are usually produced in small batches for research, the scale-up of these systems might be challenging. This is because this process is largely plagued by variations in size, shapes as well as other properties (Kim et al., 2014). It is however advocated that self-assemble nano-based vaccine systems be developed to tackle the many challenges of large-scale production protocols. Also, the process of surface modification of nano-carriers for vaccines usually involves a time-consuming, costly and complicated process of purification (Kim et al., 2014).
8: Conclusion and future perspectives
In the absence of an approved cure for the COVID-19, vaccination remains the safest and most effective way of controlling the pandemic. The advent of nanotechnology in vaccine development has sought to ensure safe and effective delivery of antigens as well as improving the immune responses of antigens as suitable adjuvants. Nano-based systems have been known over the years for their uncanny ability to increase biocompatibility, exhibit controlled as well as targeted release profiles, ensure superior drug/antigen encapsulation to mention just a few. These desirable properties have been explored in the development of COVID-19 vaccines. Nano-based strategies have precipitated the development of vaccines at ultra-rapid rates which is of huge importance in a pandemic such as this. Owing to the delicate and ravaging nature of the COVID-19, various considerations are made in the development of suitable vaccines. The structural and sub-structural proteins from the coronavirus SARS-CoV-2 play diverse roles as target sites for vaccine development. Among the four structural proteins, spike protein (S protein) of the coronavirus SARS-CoV-2 is the primary target for most preclinical and clinical researches ongoing in the development of vaccines. Various components of the S protein are exploited as antigens and about 35 out of the 47 vaccine candidates undergoing clinical trials are based on S protein. This study has provided updates on the current and advanced nanotechnological strategies employed for the development of COVID-19 vaccines. The nucleic acid vaccine candidate (mRNA-1273) by Moderna and currently in use is encapsulated in lipid nanoparticles carriers. This typifies the use of nano-based systems as effective antigen delivery systems. This review also explored several nano-based adjuvants and novel delivery devices employed in the development of COVID-19 vaccines. Also, the application of nanotechnology in the development of these vaccine candidates has created a wide range of formulations that could be delivered via alternative routes of administrations to the commonly used intra-muscular route. The use of nanotechnological strategies in the development of COVID-19 vaccines is still at the incipient stage as very few of these systems are commercially available. However, these nano-based systems are relevant tools that could play an excellent role in combating the pandemic.
References
Abdellatif, Tawfeek, Abdelfattah, El-Saber Batiha and Hetta, 2021 Abdellatif A.A.H., Tawfeek H.M., Abdelfattah A., El-Saber Batiha G., Hetta H.F. Recent updates in COVID-19 with emphasis on inhalation therapeutics: Nanostructured and targeting systems. Journal of Drug Delivery Science and Technology. 2021. ;63:102435. https://doi.org/10.1016/j.jddst.2021.102435.
Abdelmageed et al., 2020 Abdelmageed M.I., Abdelmoneim A.H., Mustafa M.I., Elfadol N.M., Murshed N.S., Shantier S.W., et al. Design of a Multiepitope-Based Peptide Vaccine against the e protein of human COVID-19: An Immunoinformatics approach. BioMed Research International. 2020;2020:doi:10.1155/2020/2683286.
Afkhami, Yao and Xing, 2016 Afkhami S., Yao Y., Xing Z. Methods and clinical development of adenovirus-vectored vaccines against mucosal pathogens. Molecular Therapy - Methods and Clinical Development. 2016;3:16030. doi:10.1038/mtm.2016.30.
Ahmad et al., 2021 Ahmad J., Hanif A., Farooqi T., Hyder F., Anwar S., Rengasamy K.R.R. Biomedicine & Pharmacotherapy Targets and strategies for vaccine development against SARS-CoV-2. Biomedicine & Pharmacotherapy. 2021;137:111254doi:10.1016/j.biopha.2021.111254.
Ahmed et al., 2021 Ahmed E., Sajjad N., Ali A., Aldakeel F.M., Mateen A., Alqahtani M.S., et al. International Immunopharmacology SARS-CoV-2 : Insight in genome structure , pathogenesis and viral receptor binding analysis – An updated review. International Immunopharmacology. 2021;95:doi:10.1016/j.intimp.2021.107493 107493.
Alexandrova, Beykov, Vassilev, Jukić and Podlipnik, 2021 Alexandrova R., Beykov P., Vassilev D., Jukić M., Podlipnik Č. The virus that shook the world: Questions and answers about SARS-CoV-2 and COVID-19. Biotechnology and Biotechnological Equipment. 2021;35(1):74–102. doi:10.1080/13102818.2020.1847683.
Alimardani, Abolmaali and Tamaddon, 2021 Alimardani V., Abolmaali S.S., Tamaddon A.M. Recent advances on nanotechnology-based strategies for prevention, diagnosis, and treatment of coronavirus infections. Journal of Nanomaterials. 2021;2021:doi:10.1155/2021/9495126.
Almazán et al., 2013 Almazán F., Dediego M.L., Sola I., Zuñiga S., Nieto-Torres J.L., Marquez-Jurado S., et al. Engineering a replication-competent, propagation-defective middle east respiratory syndrome coronavirus as a vaccine candidate. MBio. 2013;4(5):1–11. doi:10.1128/mBio.00650-13.
Alphandéry, 2020 Alphandéry E. The potential of various nanotechnologies for coronavirus diagnosis/treatment highlighted through a literature analysis. Bioconjugate Chemistry. 2020;31(8):1873–1882. doi:10.1021/acs.bioconjchem.0c00287.
Alving, Beck, Matyas and Rao, 2016 Alving C.R., Beck Z., Matyas G.R., Rao M. Liposomal adjuvants for human vaccines. Expert Opinion on Drug Delivery. 2016;13(6):807–816. doi:10.1517/17425247.2016.1151871.
An et al., 2020 An X., Martinez M.P., Rezvan A., Sefat S.R., Fathi M., Singh S., et al. Single-dose intranasal vaccination elicits systemic and mucosal immunity against SARS-CoV-2. SSRN Electronic Journal. 2020;24(9):1–44. doi:10.2139/ssrn.3751056.
Ashraf et al., 2021 Ashraf M.U., Kim Y., Kumar S., Seo D., Ashraf M., Bae Y.S. Covid-19 vaccines (revisited) and oral-mucosal vector system as a potential vaccine platform. Vaccine. 2021;9(2):1–24. doi:10.3390/vaccines9020171.
Bahloul, Lassoued and Sfar, 2014 Bahloul B., Lassoued M.A., Sfar S. A novel approach for the development and optimization of self emulsifying drug delivery system using HLB and response surface methodology : Application to feno fi brate encapsulation. International Journal of Pharmaceutics. 2014;466(1–2):341–348. doi:10.1016/j.ijpharm.2014.03.040.
Banerjee, Santra and Maiti, 2020 Banerjee A., Santra D., Maiti S. Energetics based epitope screening in SARS CoV-2 (COVID 19) spike glycoprotein by Immuno-informatic analysis aiming to a suitable vaccine development. Journal of Translational Medicine. 2020. ;18(1). https://doi.org/10.1186/s12967-020-02435-4.
Baric et al., 2016 Baric R.S., David C.S., Zhao J., Zhao J., Mangalam A.K., Channappanavar R., et al. Airway memory CD4 + T cells mediate protective immunity against emerging respiratory article airway memory CD4 + T cells mediate protective immunity against emerging respiratory coronaviruses. Immunity. 2016;44(6):1379–1391. doi:10.1016/j.immuni.2016.05.006.
Bhattacharya et al., 2016 Bhattacharya K., Mukherjee S.P., Gallud A., Burkert S.C., Bistarelli S., Bellucci S., et al. Biological interactions of carbon-based nanomaterials: From coronation to degradation. Nanomedicine: Nanotechnology, Biology, and Medicine. 2016;12(2):333–351. doi:10.1016/j.nano.2015.11.011.
Bonam et al., 2021 Bonam S.R., Kotla N.G., Bohara R.A., Rochev Y., Webster T.J., Bayry J. Potential immuno-nanomedicine strategies to fight COVID-19 like pulmonary infections. Nano Today. 2021;36:101051doi:10.1016/j.nantod.2020.101051.
Boopathi, Poma and Kolandaivel, 2020 Boopathi S., Poma A.B., Kolandaivel P. Novel 2019 coronavirus structure, mechanism of action, antiviral drug promises and rule out against its treatment. Journal of Biomolecular Structure and Dynamics. 2020;0:1–10. doi:10.1080/07391102.2020.1758788.
Campos et al., 2020 Campos E.V.R., Pereira A.E.S., De Oliveira J.L., Carvalho L.B., Guilger-Casagrande M., De Lima R., et al. How can nanotechnology help to combat COVID-19? Opportunities and urgent need. Journal of Nanobiotechnology. 2020;18(1):1–23. doi:10.1186/s12951-020-00685-4.
Chahal et al., 2016 Chahal J.S., Khan O.F., Cooper C.L., McPartlan J.S., Tsosie J.K., Tilley L.D., et al. Dendrimer-RNA nanoparticles generate protective immunity against lethal ebola, H1N1 influenza, and toxoplasma gondii challenges with a single dose. Proceedings of the National Academy of Sciences of the United States of America. 2016;113(29):E4133–E4142. doi:10.1073/pnas.1600299113.
Chang et al., 2006 Chang C.K., Sue S.C., Yu T.H., Hsieh C.M., Tsai C.K., Chiang Y.C., et al. Modular organization of SARS coronavirus nucleocapsid protein. Journal of Biomedical Science. 2006;13(1):59–72. doi:10.1007/s11373-005-9035-9.
Chao et al., 2020 Chao J.Y., Derespina K.R., Herold B.C., Goldman D.L., Aldrich M., Weingarten J., et al. Clinical characteristics and outcomes of hospitalized and critically ill children and adolescents with coronavirus disease 2019 at a tertiary care medical Center in new York City. Journal of Pediatrics. 2020;223(August):14–19.e2. doi:10.1016/j.jpeds.2020.05.006.
Chauhan et al., 2020 Chauhan G., Madou M.J., Kalra S., Chopra V., Ghosh D., Martinez-Chapa S.O. Nanotechnology for COVID-19: Therapeutics and vaccine research. ACS Nano. 2020;14(7):7760–7782. doi:10.1021/acsnano.0c04006.
Chen, Liu and Guo, 2020 Chen Y., Liu Q., Guo D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. Journal of Medical Virology. 2020;92(4):418–423.
Christensen, Smith, Andersen and Agger, 2011 Christensen D., Smith K., Andersen P., Agger E.M. Cationic liposomes as vaccine adjuvants. Expert Review of Vaccines. 2011;10(4):513–521.
Chung, Beiss, Fiering and Steinmetz, 2020 Chung Y.H., Beiss V., Fiering S.N., Steinmetz N.F. Covid-19 vaccine frontrunners and their nanotechnology design. ACS Nano. 2020;14(10):12522–12537. doi:10.1021/acsnano.0c07197.
Donato et al., 1999 Donato S.D., Grano D., Minutello M., Lecchi G., Faccini M., Agnello M. Safety and immunogenicity of MF59-adjuvanted influenza vaccine in the elderly. Vaccine. 1999;17:3094–3101.
De Soto, 2021 De Soto J.A. Evaluation of the Moderna, Pfizer/biotech, Astrazeneca/Oxford and sputnik V vaccines for Covid-19. Advance Research Journal of Medical Clinical and Science. 2021. ;07(01):408–414. https://doi.org/10.15520/arjmcs.v7i01.246.
DeDiego et al., 2007 DeDiego M.L., Álvarez E., Almazán F., Rejas M.T., Lamirande E., Roberts A., et al. A severe acute respiratory syndrome coronavirus that lacks the E gene is attenuated in vitro and in vivo. Journal of Virology. 2007;81(4):1701–1713. doi:10.1128/jvi.01467-06.
Draper and Heeney, 2010 Draper S.J., Heeney J.L. Viruses as vaccine vectors for infectious diseases and cancer. Nature Reviews Microbiology. 2010;8(1):62–73. doi:10.1038/nrmicro2240.
Erasmus et al., 2020 Erasmus J.H., Med S.T., Erasmus J.H., Khandhar A.P., Connor M.A.O., Walls A.C., et al. An alphavirus-derived replicon RNA vaccine induces SARS-CoV-2 neutralizing antibody and T cell responses in mice and nonhuman primates. Science Translational Medicine. 2020. ;12(555):1–17. https://doi.org/10.1126/scitranslmed.abc9396.
Gálvez-Barrón et al., 2021 Gálvez-Barrón C., Arroyo-Huidobro M., Miňarro A., Añaños G., Chamero A., Martín M., et al. COVID-19: Clinical presentation and prognostic factors of severe disease and mortality in the oldest-old population: A cohort study. Gerontology. 2021;1–14:doi:10.1159/000515159.
Garciá, Mancilla-Galindo, Paredes-Paredes, Tiburcio and Ávila-Vanzzini, 2021 Garciá M.M.A., Mancilla-Galindo J., Paredes-Paredes M., Tiburcio Á.Z., Ávila-Vanzzini N. Mechanisms of infection by SARS-CoV-2, inflammation and potential links with the microbiome. Future Virology. 2021;16(1):43–57. doi:10.2217/fvl-2020-0310.
Ghaffari et al., 2021 Ghaffari M., Mollazadeh-Bajestani M., Moztarzadeh F., Uludağ H., Hardy J.G., Mozafari M. An overview of the use of biomaterials, nanotechnology, and stem cells for detection and treatment of COVID-19: Towards a framework to address future global pandemics. Emergent Materials. 2021;4(1):19–34. doi:10.1007/s42247-020-00143-9.
Grifoni et al., 2020 Grifoni A., Weiskopf D., Ramirez S.I., Mateus J., Dan J.M., Moderbacher C.R., et al. Targets of T cell responses to SARS-CoV-2 coronavirus in humans with COVID-19 disease and unexposed individuals. Cell. 2020. https://doi.org/10.1016/j.cell.2020.05.015.
Hagan, 2007 Hagan D.T.O. MF59 is a safe and potent vaccine adjuvant that enhances protection against influenza virus infection. Expert Review of Vaccines. 2007. ;6(5):699–710. https://doi.org/10.1586/14760584.6.5.699.
Harrison, Lin and Wang, 2020 Harrison A.G., Lin T., Wang P. Mechanisms of SARS-CoV-2 transmission and pathogenesis. Trends in Immunology. 2020;41(12):1100–1115. doi:10.1016/j.it.2020.10.004.
Hassan et al., 2020 Hassan A.O., Kafai N.M., Dmitriev I.P., Fox J.M., Smith B.K., Harvey I.B., et al. A single-dose intranasal ChAd vaccine protects upper and lower respiratory tracts against SARS-CoV-2. Cell. 2020;183(1):169–184. doi:10.1016/j.cell.2020.08.026.
He et al., 2004 He Y., Zhou Y., Wu H., Luo B., Chen J., Li W., et al. Identification of Immunodominant sites on the spike protein of severe acute respiratory syndrome (SARS) coronavirus: Implication for developing SARS diagnostics and vaccines. The Journal of Immunology. 2004;173(6):4050–4057. doi:10.4049/jimmunol.173.6.4050.
Ho et al., 2001 Ho J.C., Hu W.H.C., Lam W.K., Chu L.W., Zheng L., Tipoe G.L., et al. The effect of aging on nasal mucociliary clearance, beat frequency and ultrastructure of respiratory cilia. In: The 6th Medical Research Conference, Hong Kong, China, 23(13–14); 2001:66.
Hobernik and Bros, 2018 Hobernik D., Bros M. DNA vaccines—How far from clinical use?. International Journal of Molecular Sciences. 2018;19(11):1–28. doi:10.3390/ijms19113605.
Hogan et al., 2001 Hogan R.J., Usherwood E.J., Zhong W., Roberts A.A., Dutton R.W., Harmsen A.G., et al. Activated antigen-specific CD8 + T cells persist in the lungs following recovery from respiratory virus infections. Journal of Immunology. 2001;166:1813–1822.
Hurst, Koetzner and Masters, 2009 Hurst K.R., Koetzner C.A., Masters P.S. Identification of in vivo-interacting domains of the murine coronavirus Nucleocapsid protein. Journal of Virology. 2009;83(14):7221–7234. doi:10.1128/jvi.00440-09.
Jackson et al., 2020 Jackson L.A., Anderson E.J., Rouphael N.G., Roberts P.C., Makhene M., Coler R.N., et al. An mRNA vaccine against SARS-CoV-2 — Preliminary report. New England Journal of Medicine. 2020;383(20):1920–1931. doi:10.1056/NEJMoa2022483.
Jackson, Kester, Casimiro, Gurunathan and DeRosa, 2020 Jackson N.A.C., Kester K.E., Casimiro D., Gurunathan S., DeRosa F. The promise of mRNA vaccines: A biotech and industrial perspective. Npj Vaccines. 2020;5(1):3–8. doi:10.1038/s41541-020-0159-8.
Jeyanathan et al., 2020 Jeyanathan M., Afkhami S., Smaill F., Miller M.S., Lichty B.D., Xing Z. - mmunological considerations for COVID-19 vaccine strategies. Nature Reviews Immunology. 2020;20(October):615–632. doi:10.1038/s41577-020-00434-6.
Jiang, Hillyer and Du, 2020 Jiang S., Hillyer C., Du L. Jiang S, Hillyer C, Du L. neutralizing antibodies against SARS- CoV-2 and other human coronaviruses. Trends in Immunology. 2020. ;41(5):355–359. https://doi.org/10.1016/j.it.2020.03.007.
Jimenez-Guardeño et al., 2015 Jimenez-Guardeño J.M., Regla-Nava J.A., Nieto-Torres J.L., DeDiego M.L., Castaño-Rodriguez C., Fernandez-Delgado R., et al. Identification of the mechanisms causing reversion to virulence in an attenuated SARS-CoV for the Design of a Genetically Stable Vaccine. PLoS Pathogens. 2015;11(10):1–36. doi:10.1371/journal.ppat.1005215.
Karpiński, Ożarowski, Mrozikiewicz and Wolski, 2021 Karpiński T.M., Ożarowski M., Mrozikiewicz A.S., Wolski H. Theranostics the 2020 race towards SARS-CoV-2 specific vaccines. Theranostics. 2021;11(4):1690–1702. doi:10.7150/thno.53691.
Keech et al., 2020 Keech C., Albert G., Cho I., Robertson A., Reed P., Neal S., et al. Phase 1–2 trial of a SARS-CoV-2 recombinant spike protein nanoparticle vaccine. New England Journal of Medicine. 2020;383(24):2320–2332. doi:10.1056/nejmoa2026920.
Kim et al., 2020 Kim E., Erdos G., Huang S., Kenniston T., Balmert S., Carey C., et al. Microneedle array delivered recombinant coronavirus vaccines: Immunogenicity and rapid translational development. eBioMedicine. 2020;55:102743.
Kim et al., 2014 Kim M.G., Park J.Y., Shon Y., Kim G., Shim G., Oh Y.K. Nanotechnology and vaccine development. Asian Journal of Pharmaceutical Sciences. 2014;9(5):227–235. doi:10.1016/j.ajps.2014.06.002.
Ksiazek et al., 2003 Ksiazek T.G., Erdman D., Goldsmith C.S., Zaki S.R., Peret T., Emery S., et al. A novel coronavirus associated with severe acute respiratory syndrome. New England Journal of Medicine. 2003;348(20):1953–1966. doi:10.1056/NEJMoa030781.
Kumar and Kumar, 2021 Kumar A., Kumar A. Mucosal and transdermal vaccine delivery strategies against COVID-19. Drug Delivery and Translational Research. 2021. ;0123456789. https://doi.org/10.1007/s13346-021-01001-9.
Kusi et al., 2011 Kusi K.A., Remarque E.J., Riasat V., Walraven V., Thomas A.W., Faber B.W., et al. Safety and immunogenicity of multi-antigen AMA1-based vaccines formulated with CoVaccine HT™ and Montanide ISA 51 in rhesus macaques. Malaria Journal. 2011;10(1):182. doi:10.1186/1475-2875-10-182.
Kuwentrai et al., 2021 Kuwentrai C., Yu J., Rong L., Zhang B.Z., Hu Y.F., Gong H.R., et al. Intradermal delivery of receptor-binding domain of SARS-CoV-2 spike protein with dissolvable microneedles to induce humoral and cellular responses in mice. Bioengineering and Translational Medicine. 2021;6(1):e10202doi:10.1002/btm2.10202.
Lehrer et al., 2018 Lehrer A.T., Wong T.A.S., Lieberman M.M., Humphreys T., Clements D.E., Bakken R.R., et al. Recombinant proteins of Zaire ebolavirus induce potent humoral and cellular immune responses and protect against live virus infection in mice. Vaccine. 2018;36(22):3090–3100. doi:10.1016/j.vaccine.2017.01.068.
Lokugamage et al., 2008 Lokugamage K.G., Yoshikawa-Iwata N., Ito N., Watts D.M., Wyde P.R., Wang N., et al. Chimeric coronavirus-like particles carrying severe acute respiratory syndrome coronavirus (SCoV) S protein protect mice against challenge with SCoV. Vaccine. 2008;26(6):797–808. doi:10.1016/j.vaccine.2007.11.092.
Lutz et al., 2017 Lutz J., Lazzaro S., Habbeddine M., Schmidt K.E., Baumhof P., Mui B.L., et al. Unmodified mRNA in LNPs constitutes a competitive technology for prophylactic vaccines. Npj Vaccines. 2017;2(1):1–9. doi:10.1038/s41541-017-0032-6.
Macera, De Angelis, Sagnelli and Coppola, 2020 Macera M., De Angelis G., Sagnelli C., Coppola N. Clinical presentation of covid-19: Case series and review of the literature. International Journal of Environmental Research and Public Health. 2020;17(14):1–11. doi:10.3390/ijerph17145062.
Machhi et al., 2021 Machhi J., Shahjin F., Das S., Patel M., Abdelmoaty M.M., Cohen J.D., et al. Nanocarrier vaccines for SARS-CoV-2. Advanced Drug Delivery Reviews. 2021;171(January):215–239. doi:10.1016/j.addr.2021.01.002.
Magnusson et al., 2018 Magnusson S.E., Altenburg A.F., Bengtsson K.L., Bosman F., de Vries R.D., Rimmelzwaan G.F., et al. Matrix-M™ adjuvant enhances immunogenicity of both protein- and modified vaccinia virus Ankara-based influenza vaccines in mice. Immunologic Research. 2018;66(2):224–233. doi:10.1007/s12026-018-8991-x.
Malabadi, Meti and Chalannavar, 2021 Malabadi R.B., Meti N.T., Chalannavar R.K. Applications of nanotechnology in vaccine development for coronavirus (SARS-CoV-2) disease (Covid-19). International Journal of Research and Scientific Innovation (IJRSI). 2021. ;8(2):191–198. Retrieved from www.rsisinternational.org.
Mason, 2020 Mason R.J. Pathogenesis of COVID-19 from a cell biology perspective. European Respiratory Journal. 2020;doi:10.1183/13993003.00607-2020.
Mba, Sharndama, Osondu-chuka and Okeke, 2021 Mba I.E., Sharndama H.C., Osondu-chuka G.O., Okeke O.P. Immunobiology and nanotherapeutics of severe acute respiratory syndrome 2 (SARS-CoV-2): A current update. Infectious Diseases. 2021;53(8):559–580. doi:10.1080/23744235.2021.1916071.
McKay et al., 2020 McKay P.F., Hu K., Blakney A.K., Samnuan K., Bouton C.R., Rogers P., et al. Self-amplifying RNA SARS-CoV-2 lipid nanoparticle vaccine induces equivalent preclinical antibody titers and viral neutralization to recovered COVID-19 patients. BioRxiv. 2020;1–14. doi:10.1101/2020.04.22.055608.
Menon et al., 2021 Menon I., Bagwe P., Gomes K.B., Bajaj L., Gala R., Uddin M.N., et al. Microneedles: A new generation vaccine delivery system. Micromachines. 2021;12(4):1–18. doi:10.3390/mi12040435.
Mueller et al., 2020 Mueller S., Stauft C.B., Kalkeri R., Koidei F., Kushnir A., Tasker S., et al. A codon-pair deoptimized live-attenuated vaccine against respiratory syncytial virus is immunogenic and efficacious in non-human primates. Vaccine. 2020;xxxx:2–7. doi:10.1016/j.vaccine.2020.02.056.
Murdin, Barreto and Plotkin, 1996 Murdin A.D., Barreto L., Plotkin S. Inactivated poliovirus vaccine: Past and present experience. Vaccine. 1996;14(8):735–746. doi:10.1016/0264-410X(95)00211-I.
Naskalska et al., 2018 Naskalska A., Dabrowska A., Nowak P., Szczepanski A., Jasik K., Milewska A., et al. Novel coronavirus-like particles targeting cells lining the respiratory tract. PLoS One. 2018;13(9):1–21. doi:10.1371/journal.pone.0203489.
Netland et al., 2010 Netland J., DeDiego M.L., Zhao J., Fett C., Álvarez E., Nieto-Torres J.L., et al. Immunization with an attenuated severe acute respiratory syndrome coronavirus deleted in E protein protects against lethal respiratory disease. Virology. 2010;399(1):120–128. doi:10.1016/j.virol.2010.01.004.
Nieto-Torres et al., 2014 Nieto-Torres J.L., DeDiego M.L., Verdiá-Báguena C., Jimenez-Guardeño J.M., Regla-Nava J.A., Fernandez-Delgado R., et al. Severe acute respiratory syndrome coronavirus envelope protein Ion Channel activity promotes virus fitness and pathogenesis. PLoS Pathogens. 2014;10(5):e1004077doi:10.1371/journal.ppat.1004077.
O'Shea, Prausnitz and Rouphael, 2021 O'Shea J., Prausnitz M.R., Rouphael N. Dissolvable microneedle patches to enable increased access to vaccines against SARS-CoV-2 and future pandemic outbreaks. Vaccine. 2021;9(4):320. doi:10.3390/vaccines9040320.
Pandey et al., 2021 Pandey A., Nikam A.N., Mutalik S.P., Fernandes G., Shreya A.B., Padya B.S., et al. Architectured therapeutic and diagnostic Nanoplatforms for combating SARS-CoV-2: Role of inorganic, organic, and radioactive materials. ACS Biomaterials Science & Engineering. 2021;7(1):31–54. doi:10.1021/acsbiomaterials.0c01243.
Pardi, Hogan, Porter and Weissman, 2018 Pardi N., Hogan M.J., Porter F.W., Weissman D. mRNA vaccines-a new era in vaccinology. Nature Reviews Drug Discovery. 2018;17(4):261–279. doi:10.1038/nrd.2017.243.
Petkar et al., 2021 Petkar K.C., Patil S.M., Chavhan S.S., Kaneko K., Sawant K.K., Kunda N.K., et al. An overview of nanocarrier-based adjuvants for vaccine delivery. Pharmaceutics. 2021;13(4):1–29. doi:10.3390/pharmaceutics13040455.
Petsch et al., 2012 Petsch B., Schnee M., Vogel A.B., Lange E., Hoffmann B., Voss D., et al. Protective efficacy of in vitro synthesized, specific mRNA vaccines against influenza a virus infection. Nature Biotechnology. 2012;30(12):1210–1216. doi:10.1038/nbt.2436.
Plotkin, Orenstein, Offit, & Edwards, 2017 Plotkin S.A., Orenstein, W.A., Offit P.A., Edwards K.M. Vaccines E-book. Elsevier Health Sciences; 2017.
Polack et al., 2020 Polack F., Thomas S., Kitchin N., Absalon J., Gurtman A., Lockhart S., et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. The New England Journal of Medicine. 2020. ;10:1–13. https://doi.org/10.1056/NEJMoa2034577.
Poon et al., 2018 Poon C., Gallo J., Joo J., Chang T., Bañobre-López M., Chung E.J. Hybrid, metal oxide-peptide amphiphile micelles for molecular magnetic resonance imaging of atherosclerosis. Journal of Nanobiotechnology. 2018;16(1):1–11. doi:10.1186/s12951-018-0420-8.
Qian et al., 2012 Qian Z., Travanty E.A., Oko L., Edeen K., Berglund A., Wang J., et al. Innate immune response of human alveolar type II cells infected with severe acute respiratory syndrome – Coronavirus. American Journal of Respiratory Cell and Molecular Biology. 2012;10(22):22–26. doi:10.1165/rcmb.2012-0339OC.
Rahman et al., 2020 Rahman H.S., Aziz M.S., Hussein R.H., Othman H.H., Salih Omer S.H., Khalid E.S., et al. The transmission modes and sources of COVID-19: A systematic review. International Journal of Surgery Open. 2020;26:125–136. doi:10.1016/j.ijso.2020.08.017.
Rao et al., 2020 Rao L., Xia S., Xu W., Tian R., Yu G., Gu C., et al. Decoy nanoparticles protect against COVID-19 by concurrently adsorbing viruses and inflammatory cytokines. Proceedings of the National Academy of Sciences of the United States of America. 2020;117(44):27141–27147. doi:10.1073/pnas.2014352117.
Raoufi, Bahramimeimandi, Salehi-Shadkami, Chaosri and Mozafari, 2021 Raoufi E., Bahramimeimandi B., Salehi-Shadkami M., Chaosri P., Mozafari M.R. Methodical design of viral vaccines based on avant-Garde nanocarriers: A multi-domain narrative review. Biomedicine. 2021;9(5):1–20. doi:10.3390/biomedicines9050520.
Reza-Zaldıvar, 2021 Reza-Zaldıvar E.E., et al. Infection mechanism of SARS-COV-2 and its implication on the nervous system. Frontiers in Immunology. 2021;11:621735doi:10.3389/fimmu.2020.621735 In press.
Ruiz-Hitzky et al., 2020 Ruiz-Hitzky E., Darder M., Wicklein B., Ruiz-Garcia C., Martín-Sampedro R., del Real G., et al. Nanotechnology responses to COVID-19. Advanced Healthcare Materials. 2020;9(19):1–26. doi:10.1002/adhm.202000979.
Sanclemente-Alaman et al., 2020 Sanclemente-Alaman I., Moreno-Jiménez L., Benito-Martín M.S., Canales-Aguirre A., Matías-Guiu J.A., Matías-Guiu J., et al. Experimental models for the study of central nervous system infection by SARS-CoV-2. Frontiers in Immunology. 2020;11(8):1–12. doi:10.3389/fimmu.2020.02163.
Sarkesh et al., 2020 Sarkesh A., Sorkhabi A.D., Sheykhsaran E., Alinezhad F., Mohammadzadeh N., Hemmat N., et al. Extrapulmonary clinical manifestations in COVID-19 patients. American Journal of Tropical Medicine and Hygiene. 2020;103(5):1783–1796. doi:10.4269/ajtmh.20-0986.
Schnee et al., 2016 Schnee M., Vogel A.B., Voss D., Petsch B., Baumhof P., Kramps T., et al. An mRNA vaccine encoding rabies virus glycoprotein induces protection against lethal infection in mice and correlates of protection in adult and newborn pigs. PLoS Neglected Tropical Diseases. 2016;10(6):1–20. doi:10.1371/journal.pntd.0004746.
Shin et al., 2020 Shin M.D., Shukla S., Chung Y.H., Beiss V., Chan S.K., Ortega-Rivera O.A., et al. COVID-19 vaccine development and a potential nanomaterial path forward. Nature Nanotechnology. 2020;15(8):646–655. doi:10.1038/s41565-020-0737-y.
Soleimanpour and Yaghoubi, 2021 Soleimanpour S., Yaghoubi A. COVID-19 vaccine: Where are we now and where should we go? Expert Review of Vaccines: Taylor and Francis Ltd.; 2021.doi:10.1080/14760584.2021.1875824.
Strizova, Smetanova, Bartunkova and Milota, 2021 Strizova Z., Smetanova J., Bartunkova J., Milota T. Principles and challenges in anti-COVID-19 vaccine development. International Archives of Allergy and Immunology. 2021;182(4):339–349. doi:10.1159/000514225.
Suganya, Divya and Parani, 2021 Suganya S., Divya S., Parani M. Severe acute respiratory syndrome-coronavirus-2: Current advances in therapeutic targets and drug development. Reviews in Medical Virology. 2021;doi:10.1002/rmv.2174.
Tai et al., 2020 Tai W., He L., Zhang X., Pu J., Voronin D., Jiang S., et al. Characterization of the receptor-binding domain (RBD) of 2019 novel coronavirus: Implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cellular and molecular immunology. 2020;17(6):613–620. doi:10.1038/s41423-020-0400-4.
Tran et al., 2020 Tran K.T.M., Gavitt T.D., Farrell N.J., Curry E.J., Mara A.B., Patel A., et al. Transdermal microneedles for the programmable burst release of multiple vaccine payloads. Nature Biomedical Engineering. 2020;doi:10.1038/s41551-020-00650-4.
Turoňová et al., 2020 Turoňová B., Sikora M., Schürmann C., Hagen W.J.H., Welsch S., Blanc F.E.C., et al. In situ structural analysis of SARS-CoV-2 spike reveals flexibility mediated by three hinges. Science. 2020;370(6513):203–208. doi:10.1126/science.abd5223.
van Doremalen et al., 2020 van Doremalen N., Lambe T., Spencer A., Belij-Rammerstorfer S., Purushotham J.N., Port J.R., et al. ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature. 2020;586(7830):578–582. doi:10.1038/s41586-020-2608-y.
Vellozzi et al., 2009 Vellozzi C., Burwen D.R., Dobardzic A., Ball R., Walton K., Haber P. Safety of trivalent inactivated influenza vaccines in adults: Background for pandemic influenza vaccine safety monitoring. Vaccine. 2009;27(15):2114–2120. doi:10.1016/j.vaccine.2009.01.125.
Wan, Shang, Graham, Baric and Li, 2020 Wan Y., Shang J., Graham R., Baric R.S., Li F. Receptor recognition by the novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS coronavirus. Journal of Virology. 2020;94(7):doi:10.1128/jvi.00127-20.
Wang et al., 2020 Wang D., Hu B., Hu C., Zhu F., Liu X., Zhang J., et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA : The Journal of the American Medical Association. 2020;323(11):1061–1069. doi:10.1001/jama.2020.1585.
Wang, Peng, Xu, Cui and Williams, 2020 Wang J., Peng Y., Xu H., Cui Z., Williams R.O. The COVID-19 vaccine race: Challenges and opportunities in vaccine formulation. AAPS PharmSciTech. 2020;21(6):225. doi:10.1208/s12249-020-01744-7.
Wang et al., 2020 Wang M.Y., Zhao R., Gao L.J., Gao X.F., Wang D.P., Cao J.M. SARS-CoV-2: Structure, biology, and structure-based therapeutics development. Frontiers in Cellular and Infection Microbiology. 2020;doi:10.3389/fcimb.2020.587269.
WHO, 2020 WHO. Draft landscape of COVID-19 candidate vaccines- November 3. World Heal. Organ; 2020.
World Health Organization, 2021 World Health Organization. Status of COVID-19 vaccines within WHO EUL/PQ evaluation process (20 January 2021). World Health Organization; 2021. (February), 2. Retrieved from https://extranet.who.int/pqweb/sites/default/files/documents/Status_COVID_VAX_20Jan2021_v2.pdf.
Wu and McGoogan, 2020 Wu Z., McGoogan J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72314 cases from the Chinese Center for Disease Control and Prevention. JAMA : The Journal of the American Medical Association. 2020;323(13):1239–1242. doi:10.1001/jama.2020.2648.
Wu et al., 2020 Wu F., Wang A., Liu M., Wang Q., Chen J., Xia S., et al. Neutralizing antibody responses to SARS-CoV-2 in a COVID-19 recovered patient cohort and their implications. SSRN Electronic Journal. 2020;doi:10.2139/ssrn.3566211.
Xia et al., 2020 Xia S., Zhu Y., Liu M., Lan Q., Xu W., Wu Y., et al. Fusion mechanism of 2019-nCoV and fusion inhibitors targeting HR1 domain in spike protein. Cellular and molecular immunology. 2020. https://doi.org/10.1038/s41423-020-0374-2.
Yang, 2021 Yang D. Application of nanotechnology in the COVID-19 pandemic. International Journal of Nanomedicine. 2021;16:623–649. doi:10.2147/IJN.S296383.
Young, Thone and Jik, 2021 Young J., Thone M.N., Jik Y. COVID-19 vaccines : The status and perspectives in delivery points of view. Advanced Drug Delivery Reviews. 2021;170:1–25. doi:10.1016/j.addr.2020.12.011.
Zarandi, Zinatizadeh, Zinatizadeh, Yousefi and Rezaei, 2021 Zarandi P.K., Zinatizadeh M.R., Zinatizadeh M., Yousefi M.H., Rezaei N. SARS-CoV-2: From the pathogenesis to potential anti-viral treatments. Biomedicine & Pharmacotherapy. 2021;137:111352doi:10.1016/j.biopha.2021.111352.
Zhang et al., 2020 Zhang N.N., Li X.F., Deng Y.Q., Zhao H., Huang Y.J., Yang G., et al. A thermostable mRNA vaccine against COVID-19. Cell. 2020;182(5):1271–1283. doi:10.1016/j.cell.2020.07.024.
Zhang et al., 2017 Zhang D.Y., Zheng Y., Tan C.P., Sun J.H., Zhang W., Ji L.N., et al. Graphene oxide decorated with Ru(II)-polyethylene glycol complex for lysosome-targeted imaging and photodynamic/Photothermal therapy. ACS Applied Materials and Interfaces. 2017;9(8):6761–6771. doi:10.1021/acsami.6b13808.
Zhao et al., 2014 Zhao L., Seth A., Wibowo N., Zhao C.-X., Mitter N., Yu C., et al. Nanoparticle vaccines. Vaccine. 2014;32(3):327–337. doi:10.1016/j.vaccine.2013.11.069.
Zhao et al., 2020 Zhao J., Yuan Q., Wang H., Liu W., Liao X., Su Y., et al. Antibody responses to SARS-CoV-2 in patients with novel coronavirus disease 2019. Clinical Infectious Diseases. 2020;71(16):2027–2034. doi:10.1093/cid/ciaa344.
Zhou et al., 2020 Zhou P., Yang X.-L., Wang X.-G., Hu B., Zhang L., Zhang W., et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270–273. doi:10.1038/s41586-020-2012-7.