Introduction
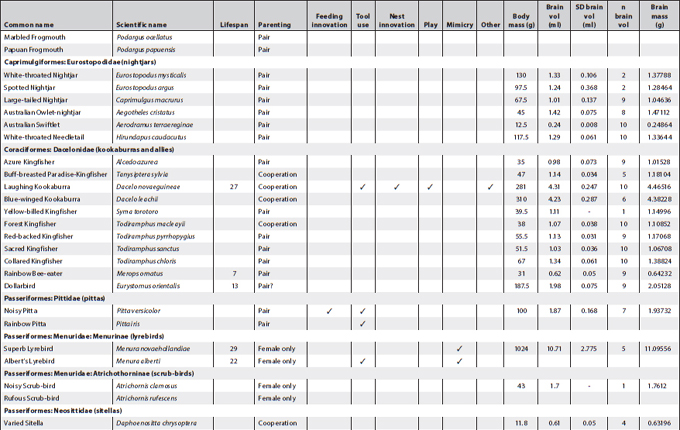
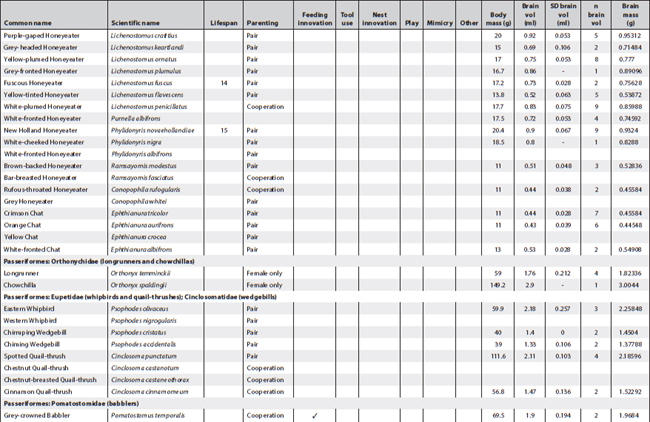
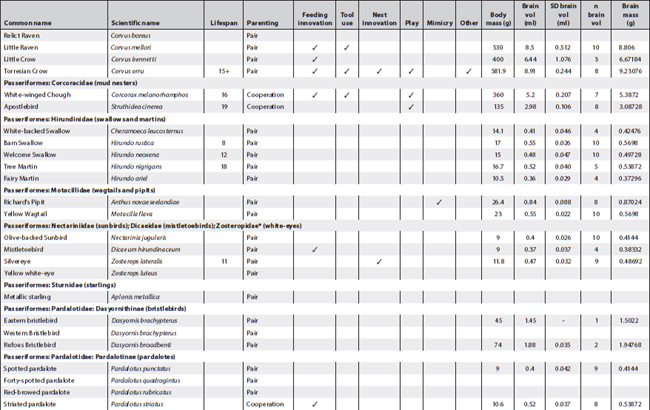
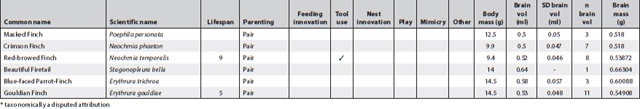
This appendix presents all known information about cognition and important related data of Australian landbirds. Where known, it provides lifespans, breeding arrangements (raising youngsters alone, in pairs or cooperatively), known behaviour/special skills suggesting a cognitive dimension, and biological data on brain and body size per species. It is a work in progress as future research efforts will no doubt fill in some of the gaps.
The species and families listed here are related to the emphasis in the book, hence do not contain waterbirds and shorebirds. Orders and families have been organised roughly in the same sequence as can be found in the widely available Field Guide to the Birds of Australia, by Simpson & Day (any edition), with the exception that pardalotes and finches are placed last instead of corvids.
The information about breeding arrangements relies heavily on A. Cockburn’s supplementary material to his 2006 paper, and for brain measurements on the paper by Franklin et al. (2014). Both papers are listed in the reference section. I want to thank Don Franklin for his generosity in sending the material to me prior to publication so that I would have time to incorporate some of his and his colleagues’ findings in this book (discussed in Chapter 2). The data reproduced here are confined to the raw data (actual weights and volume measures of each bird and of its brain) and do not include the statistical transformations. These can be ascertained from their paper. However, it is possible from the data presented here to undertake calculations about relative brain weight in addition to those provided in Franklin et al. (2014).
Of all columns, the lifespan column is, as always, the least satisfactory – even when figures are presented they ought to be viewed with some caution. For parrots and cockatoos, the main source used was: Brouwer et al. (2000), published in the International Zoo Yearbook (for full details see the reference section). The concept of lifespan is rather problematic in the sense that very few animals can live out their full lifespan in the wild and apart from banding records, which rarely start at fledging, it is almost impossible to ascertain. Still, over many generations of banding it is possible to gain some idea of maximum lifespan of a species. However, it is difficult to arrive at an average lifespan for a species even under ideal conditions. For instance, mortality rates prior to sexual maturity may be as high as 80% in avian species. If these are included, average lifespan is very low indeed and misrepresents the 20% who do make it to adulthood and may live out their lifespan in full. The matter is further complicated by the fact that the most convenient set of data may derive from birds in captivity; if in a zoo, record keeping is more reliable but stress levels for the animals may be high and may lead to much shorter lifespans. Moreover, the zoo records I gleaned for psittacine species represent minimum lifespans, meaning that birds might have been acquired at an unknown adult age but eventually died in the zoo. The figures presented here in cases where any information was available also derive from a variety of other sources such as banding data for which HANZAB was used (also listed in the reference section), personal communication, personal experience and on fact sheets published by governmental agencies when they included information on lifespan, sometimes also called life expectancy or longevity. The Australian Bird Study Association also has a website (http://www.absa.asn.au) listing some species. That site also relies on banding data. The italicised numbers in this column refer solely to captivity data and sometimes are widely divergent because, while most captivity data stem from zoos, others are derived from private ownership.
The categories placed in between lifespan and brain and body measurements follow the chapter headings of the book and, in very broad outlines, indicate whether any observations exist suggesting any known cognitive dimension or indeed complex cognitive behaviour in Australian native birds. While there are many blanks, this appendix suggests certain clusters. In some cases, as in the bowerbirds, the white-winged choughs and the Australian magpies, nearly all boxes are ticked. This may be an indication that they are unique but could also mean that these species have been studied more extensively than others and that we may find even more extraordinary birds in future. In the case of pigeons and doves, for instance, we still have far too little information. Judging by the many experiments conducted with Columba livia (the feral or rock pigeon) one may surmise that there might be a few surprises in store if someone started testing any of the Australian species.
The data published foremost by Cockburn and Franklin have enabled me to attempt here, for the first time, to take stock of the behaviour of Australian landbirds and to expose features of that behaviour that may indicate adaptability and flexibility.