1
Life in soil (e.g. active and resting of microbial species and its distribution within the soil matrix) is the product of direct and indirect interactions between highly variable physical and chemical matrices with associated equally variable biological communities composed of essentially all life forms. Soil biological communities are sustained across an intriguingly interesting span of chemical and physical conditions, especially when compared with the variability of the comparable properties of the environment supporting humans. Unfortunately for the scientists seeking a clear and accurate understanding of the identity and functions of soil microbes, the distribution of the microbes within the soil physical structure is not necessarily readily accessible. The location in soil most important for the function of soil microbes is not the macrosystem clearly accessible for scientific study but rather the microbial community which is impacted most by the properties of the microsites of a few cubic microns wherein they occur and function. Therefore, the primary goal of this introductory chapter of Soil Microbiology is to examine the general properties of soil and their impact on the associated microbial activity. To gain an appreciation of the complexity of the soil microbe's habitat, the overall physical and chemical properties of soil particulates controlling the development of soil communities will be introduced first.
Defining the world of the soil microbes: Microbes can clearly be studied in isolation from their native habitat, i.e. in soil samples of limited size or pure cultures of soil microbes incubated in the laboratory. Indeed, there is much still to be learned by such studies, but a full appreciation of microbes and their contributions to the sustainability of ecosystems can only be fully gained, or perhaps appreciated, by examining their capacity to function within their native habitat. Thus, we must initiate our examination of microbes in soil systems with a consideration of the basic properties of the soil environment itself. As will be demonstrated by the topics presented in this text, life is difficult for the microorganisms growing within the microsites comprising their soil habitat – thereby making “life” for the microbiologist attempting to understand the soil microbial community just as difficult, if not more.
Developing a clear understanding of the dynamics of soil microbial communities presents a formidable practical and conceptual challenge. The practical challenge is that due to the nature of soil itself, most of our endeavors to study the soil microbes must be conducted in the laboratory under less than natural conditions. The conceptual challenge results from the fact that many of the assumptions that we would make regarding the properties of that portion of the soil where the microbe grows are false. We see the general properties of a soil system such as a soil with a plentiful oxygen supply, a near neutral pH, and optimal moisture when in reality the microbes may be actually growing in a vastly different situation, perhaps in an acidic environment, in the absence of molecular oxygen, under water‐limited conditions (e.g. arid or flooded soil). We, the observers, see the macro‐world that we inhabit – the microbe functions within and is controlled by a few cubic micrometers of our world. Thus, the concept of the soil properties controlling microbial development and function can be, and most certainly are, vastly different from our picture of the soil system.
The soil ecosystem is the product of intricate interactions between a physical and chemical matrix of highly variable composition and biological communities composed of essentially all life forms. It is probably realistic to assume that a reasonable understanding of the extreme variability of microsites within the soil is a more modern development. Thus, a first task in studying soil microbial community development and function in soil systems is to expand our concept of the limits of physical and chemical conditions within which microbial life can exist. Thus, it is necessary to consider that the properties of the microbial world are variable within a range that extends far beyond that which is comfortable for us. Microbial life is sustainable across nearly the entire range of chemical and physical conditions existent on planet Earth.
A further limitation impacting the validity of our picture of how microbes function in soil is the fact that most of the data we have collected that is descriptive of their function are derived from laboratory‐based studies examining individual microbial species or strains. Our ability to study the microbes directly in their “home” is limited at best. This simplification of soil physical, chemical, or biological properties, from several to perhaps a single function, has allowed for a reduction in the complexity of the system of study and of data interpretation. But at what cost – reduced understanding of the real functional complexity of the community? In part, to circumvent this limitation, more recently, use of consortia of bacteria and/or fungi for study of more complex processes, such as decomposition of xenobiotic compounds or changes in the diversity of the microbial community, has become the norm. These latter cultures exhibit some of the more complex interactions reflective of the soil ecosystem. Use of defined cultures or even the simple mixtures of microbes making up consortia is generally justified by the conclusion that it is necessary to attain maximum control of experimental variables in order to elucidate clearly the processes of interest.
As an appreciation of the complexities of soils and their microbial populations is developed, the inadequacy of experiments using axenic cultures (that is, cultures composed of a single species or strain of soil microbes) or mixed cultures composed of a small number of microbial strains (that is, a consortia of soil microbes) to evaluate the world of the soil microbes becomes obvious. Generally, the basic properties of the biological processes measured in laboratory culture defy extrapolation to the more complex soil environment. Not only are stresses of the native soil system unmatched with defined culture conditions, but the microbes themselves change phenotypically and even genotypically in response to the laboratory growth conditions. Although the microbes studied may have been isolated from a sample of a soil, their metabolic capabilities and capacity to respond to their physical and chemical environment, as well as the presence of other microbes, may differ meaningfully from the strains existent in the original soil sample. That is, a microbial variant with appropriate properties for optimal growth in test tubes is usually selected either spontaneously (as a result of laboratory culture methods) or through genetic manipulation of the microbial isolates. The former selection results from the rich genetic variations within the genome of each individual bacterial species. It is easy conceptually to assume that each bacterial species present in soil consists of a group of individuals with essentially identical genomes. This viewpoint is far from reality. Bacterial species have similar traits and genetic composition, but their genomes contain many genetic variations (mutations).
Therefore, when members of a previously soil‐resident bacterial species are selected for culture in the laboratory, mutant strains that previously existed in soil as a minuscule portion of the soil population of the species may be better able to grow under the laboratory conditions of the test tube than is the majority strain occurring in the soil community. Thus, a genetic variant becomes the dominant strain studied in the laboratory. Further selection of spontaneously occurring variants may – or more likely does – occur in the laboratory culture. As a result, data collected from experiments with isolated microbial species or strains frequently only explain or mimic the processes occurring in soil in part. Therefore, it is reasonable to conclude that soil microbiologists must expand the purview of experimental design to include the complexities – controllable and otherwise – of the total soil ecosystem. Therefore, the realm of soil microbiology must be defined to include both an understanding of the properties of the microbes themselves along with an appreciation of the impact of variability of the soil environment on these traits. Soil microbial ecology must involve an evaluation of the behavior of organisms in their native habitats. Even the simplification of microbial community dynamics to those of a simple consortium is problematic, when we understand that the microbes growing in soil, rather than functioning within a colony or even a micro‐colony, are more likely part of a biofilm that consists of several species or strains of bacteria with a variety of metabolic or physiological capabilities.
Are soil microbes really that important? To the purist, elucidation of the principles of soil microbiology is immensely interesting and fully justified strictly by the information generated relating to the basic ecological interactions occurring in soil systems. Just the fact that a single gram of soil can contain thousands of different microbial species is sufficient to pique our interest. Yet, due to societal management of terrestrial ecosystems, our concern for soil biological processes reaches far beyond just the realm of basic science. For example, from the world of environmental science, our myopic exploitation of natural ecosystems has resulted in situations where the functioning of resident microbial populations is precluded or severely limited by mismanagement. Spills of toxic organic chemicals result in scarred landscapes. Similarly, products of metal processing have resulted in sites resembling moonscapes (Figure 1.1a). Reclamation of such sites is attempted (e.g. Figure 1.1b), but for attainment of such objectives, management plans must be developed that allow for establishment of essential soil biological processes in soils containing extreme levels of toxic metals or a variety of organic toxicants. For the soils adjacent to a zinc smelter, attainment of improved microbial function required amendment of the soil with an organic matter source (sewage biosolids), liming the soil to raise the soil pH to above 7.0 (to immobilize the toxic metals), and planting the soil with a strain of grass that is capable of growing in soils with elevated metals loadings in order to stabilize the soil on the mountain slopes and reduce the soil erosion. Actually, the contributions of the grass to the microbial function extend far beyond just stabilizing the physical environment. By converting solar energy to plant biomass (photosynthesis), the plants provide the primary microbial energy source for the soil microbes.
In comparison with these obviously detrimental situations, development of soils for agricultural production may also reduce overall soil quality. Tillage initiates processes that may eventually reduce productivity of the soils. Loss of desirable soil structure and reduction of soil organic matter reserves are generally associated with implementation of intensive agricultural practices. Destruction of soil structure increases the bioavailability of native soil organic matter, a major microbial carbon and energy source. Conversion of the soil organic carbon to carbon dioxide reduces the soil productivity and sustainability, thereby reducing soil quality and increasing the production of a greenhouse gas.
A dramatic example of augmented soil organic matter decomposition due to cultivation of a previously undisturbed soil system is provided by observation of changes in soil properties resulting from cropping of peats. Development of these soils for agriculture initially required draining the water, resulting in conversion of the anoxic conditions to an aerobic state. This conversion of the generally anoxic swampy soil to one where molecular oxygen is no longer limiting resulted in stimulation of the aerobic heterotrophic bacteria. The microbes oxidized the plant debris that was accumulated under the swampy conditions prior to site drainage to carbon dioxide, mineral nutrients (e.g. mineral phosphorus and nitrogen), and water, resulting in loss of soil mass. The loss of the organic matter resulted in subsidence of the soil surface (Figure 1.2).

Figure 1.1 Metal‐impacted site in Palmerton, PA (USA). (a) Site without reclamation management. (b) Site following amendment of soil surface with a mixture of sludge, fly ash, lime, and grass seed.

Figure 1.2 Subsidence pole located in the Everglades Agricultural Area of South Florida (USA) showing loss of soil elevation between 1924 and 1979.
In each of these examples – contamination of soil with metals, agricultural cultivation of the soil, or draining of swamps for agricultural production – the physical structure of the soil, its chemical properties, and the function of the biological community have been altered in a manner that reduces the quality of the soil system. Soil is a renewable resource, but the time frame for restoration of a badly damaged soil system may extend over several decades or even centuries. Thus, in the short run, soil must be managed with the philosophy that it is an exhaustible resource.
Proper soil stewardship cannot be achieved without a full appreciation of soil microbiological processes, for the totality of existence of a soil ecosystem relies upon sustaining a functional soil microbial community. To develop appropriate management plans for restoration of damaged systems or for maintenance of ecosystem productivity, a clear understanding of in situ biological processes is necessary. This cannot be fully accomplished by using data generated from the observation of bacterial populations growing at an unnatural rate under the optimized conditions of the laboratory culture flask. Realistic estimates of metabolic activity and the kinetics of the process in the microbe's native environment must be developed. Procedures necessary for maintenance of the soil chemical and physical factors controlling expression of microbial activity in a balanced state are needed to ensure the long‐term survival of critical populations for ecosystem remediation and sustainability. Such factors as temperature, pH, moisture, surface area, osmotic strength, and presence of predators all interact to select for the development and survival of a stable microbial community necessary for long‐term ecosystem function.
The challenge: Once it is understood that reality for soil microbes exists in their natural habitat, the primary challenge becomes defining that habitat. For the soil microbiologist, this is not an easy task. Microbes flourish in temperature‐limited, native systems, under such extreme conditions as experienced in hot spring‐impacted soils or in the cold desert soils of Antarctica. Highly productive soils may become biologically impoverished due to the impact of anthropogenically produced situations, such as those resulting from acid mine drainage or spills of xenobiotic organic compounds. Thus, it becomes clear that the soil ecosystem can be spoken of in general terms, but a true encounter with soil ecology must include an assessment of the length and breadth of these highly variable systems.
This assessment must start with differentiation of the entity to be studied. Soil microbiologists must realize that they are studying specific soil types, not simply “soil.” The commonly heard statement among soil microbiologists, “Soil was used in our experiments,” would sound as strange to a soil scientist as would “A bacterium was studied” would to a biologist. Currently, the US soil classification system divides world soils into 12 orders (based on soil properties that reflect soil development). Examples of soil orders are aridisols (dry soils), andisols (volcanic soils), gelisols (permafrost‐impacted soils), histosols (organic soils), inceptisols (embrionic soils), spodosols (typical of forest systems), and vertisols (characterized by soils with high‐swelling clay contents). These orders are subdivided into suborders, great groups, subgroups, families, and series. Currently, over 15 000 soil series have been identified. (See Brady and Weil 2017 for further discussion of soil taxonomy.) This situation of defining the specific soil unit of study is further complicated by the fact that a given field site may contain one or more soil series. Furthermore, biological activity within the same soil series varies with such ecosystem depending on properties as plant type and distribution, seasonal climatic variation, and management practices – current and historic. Yet, the properties of the soil type underlying an ecosystem as defined and impacted by the aboveground communities must be assessed and understood.
1.1 Soil as an Ecosystem
The properties of a soil system are not simply a summation of the general properties of its individual physical, biological, and chemical components. They are the result of the individual properties as modified by their interaction with each other as well as their distribution within the soil matrix. These interactions result in a soil ecosystem that is truly much greater than just the sum of its parts. For example, association of the soil mineral and organic materials in soil aggregates results in co‐occurrence of anoxic and aerobic biological processes within a few millimeters or less of each other in the surface horizon of a soil permeated by our oxygen atmosphere. As will be elucidated in Chapter 2, even limited growth of a pioneer population of microbial cells can result in modification of the microbe's environment sufficiently to enhance future growth and community development. Thus, a soil ecosystem could be said to be evolving toward an optimized ecological state dictated by the interactions of each of the soil components.
Therefore, any analysis of the structure of soil systems and the principles describing their function must commence with an evaluation of (i) the soil physical components, (ii) their assembly into the physical whole, and (iii) the implications of this structure on soil biological processes. The study must then proceed to a determination of the feedback mechanisms involved with living populations interacting with their physical environment. This analysis is necessary to develop fully the principles or maxims describing microbial interactions in soil and their importance to that portion of the system most commonly regarded esthetically – the aboveground plant and animal community.
1.1.1 Soil System Function
A soil's classification could be based on an understanding of the applicable physical, chemical, and biological properties of soil that make each soil unique. For example, soil definition could be based on our understanding of the impacts of plant community development on soil ecosystem properties whereas others studying the soil system could emphasize the variation in general soil properties, emphasizing an appreciation of the foundational role of soils in societal development, function, and sustainability. Indeed, societies can rise and fall based on the capability of their soil to provide ecosystem services (French 2010). Since soil microbes are key participants in the provision of these services, societies are truly built upon the function, sustainability, and diversity of the soil microbial community. To fully appreciate the intricacy of this soil microbial community, our more general image of the material that we walk on, build our infrastructure on and depend upon for food and fiber production must be enlarged to include a picture of the microscopic world in which soil microorganisms live and function.
The soil physical structure and soil microbes: Most soils are classified as mineral soils – that is, soils that are composed primarily of mineral matter and whose physical properties are controlled by the proportions of the mineral particulates contained therein. These soils generally contain from less than 1% to approximately 4% organic matter, but they may consist of as much as 20% soil organic matter. Additionally, an organic layer up to 30 cm in depth may be found on the soil surface (e.g. a forest litter layer). These soils are to be contrasted with the much less commonly encountered organic soils (histosols). Organic soils contain more than 20% organic matter as exemplified by peats and mucks.
The most obvious components of mineral soils are sand, silt, and clay, each of which is defined by its physical dimensions (Table 1.1). Note that this separation of soil particles is based on particle size only. For example, the clay fraction is strictly defined as having a mean diameter of less than 2 μm. Therefore, this fraction will contain clay minerals plus other soil components, which are not necessarily clay minerals but are of comparable size. A soil is classified into a specific textural category by the quantities of the mineral separates that it contains (Figure 1.3)
Table 1.1 Size distribution of soil particle classes
Source: USDA Public Domain.
|
Particle class |
Subclass |
Mean diameter (mm) |
|
Sand |
Very coarse sand |
2.00–1.0 |
|
Coarse sand |
1.0–0.5 |
|
|
Medium sand |
0.5–0.25 |
|
|
Fine sand |
0.25–0.1 |
|
|
Very fine sand |
0.1–0.05 |
|
|
Silt |
0.05–0.002 |
|
|
Clay |
<0.002 |
This disposition of soil by textural classification is useful for characterizing a particular soil of interest (either for field or laboratory experiments), but it is in reality the soil structure – that is, how these individual components are assembled – that best defines the habitats of soil microorganisms. Two soils may have nearly identical textural compositions yet exhibit extremely contrasting microbial activities. For example, microbial properties of a silt loam in an area flooded with acid mine drainage waters would reflect the prevailing stresses of oxygen deprivation and extremes of acidity, whereas a silt loam from a well‐maintained agricultural site would be free of such limitations. Furthermore, a soil with a well‐developed aggregate structure in contrast to one where the structure has been lost due to intensive cultivation would be more conducive to microbial community development.
Clearly, a soil classification system is required that is descriptive of in situ ecological influences on biological properties. This information is provided by soil classification systems, such as those of the United States Department of Agriculture (USDA) or the Food and Agriculture Organization of the United Nations (FAO). Within these taxonomic systems, soils are grouped by field and laboratory properties, among which are soil texture, moisture, pH, temperature regimes, and horizon development.
Soil profile development: An appreciation of soil horizon development and its effect on soil microbial activity is necessary to develop a more complete picture of the impact of soil biological processes on the properties of an ecosystem. A soil horizon is a layer of soil approximately parallel to the land surface and differing from adjacent layers physically, chemically, and biologically, or in characteristics such as color, structure, texture, consistency, biotic populations, and pH. Soil horizonal development is clearly a product of current ecosystem properties as well as of site history. Each soil subgroup description includes an analysis of horizonal structure. Microbiological properties of a specific horizon may be studied in isolation from other portions of the soil profile. In such studies, the designated portion of the soil profile may be defined as an ecosystem in itself.
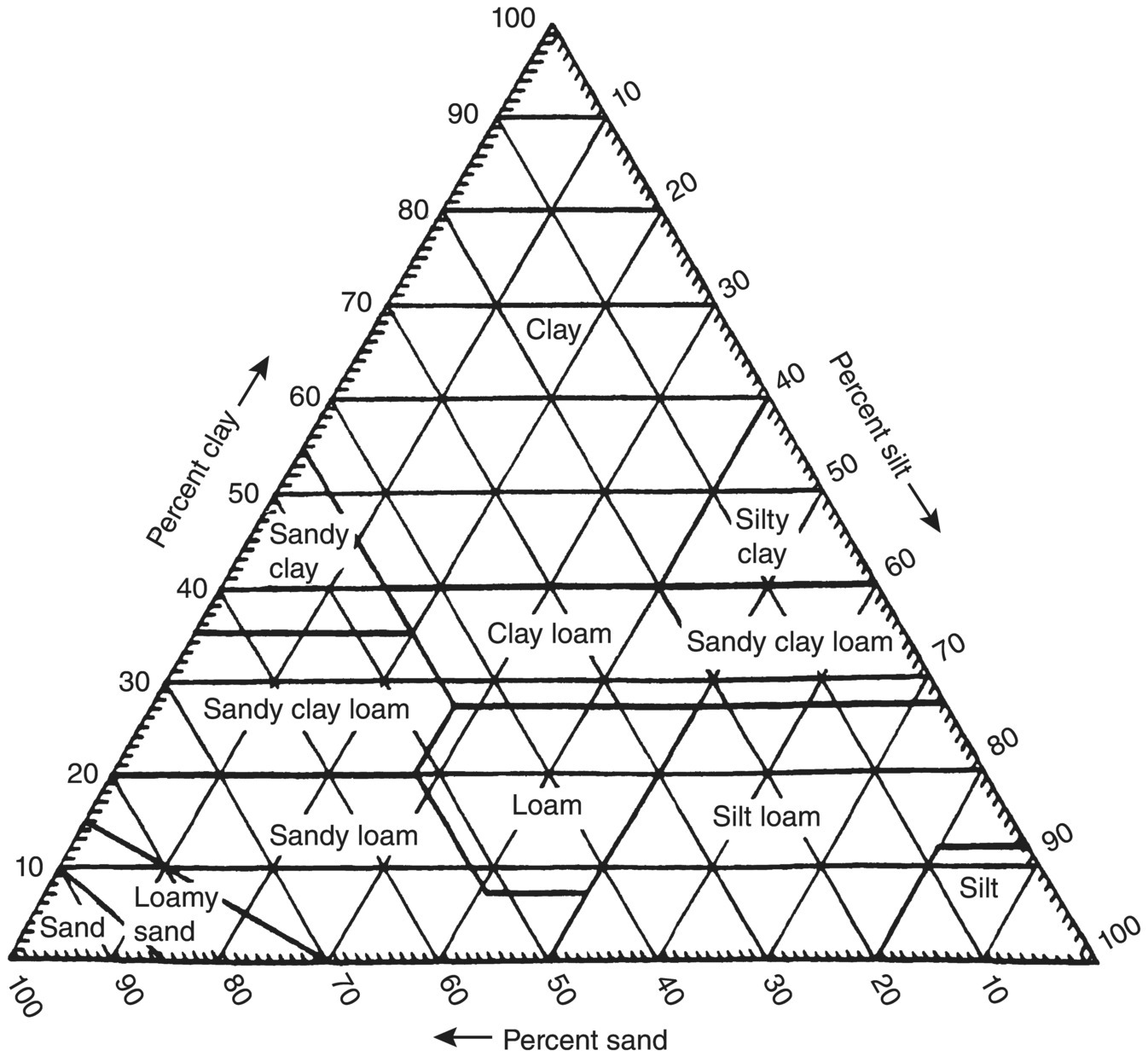
Figure 1.3 Designation of soil textural classes by principal mineral component analysis.
Source: USDA Public Domain.
The most commonly studied of soil horizons in microbiological research are the O, A, and B horizons, although as a result of complications of groundwater pollution, considerable research effort is being expended to understand biological processes occurring in deeper‐lying aquifer materials. O horizons are dominated by organic material and are exemplified by forest litter layers. The A horizons are mineral layers formed on the soil surface or below the O horizon. These regions are characterized by accumulations of organic materials intimately associated with soil mineral matter. In many situations, these organic materials are a product of plant community development in native, unmanaged ecosystems. A horizons are usually spoken of as surface soils. Colloidal organic matter concentrations tend to be maximized in this portion of the mineral soil profile. B horizons are usually formed below an O or A horizon and are dominated by (i) carbonates, gypsum, or silica, alone or in combination; (ii) evidence of removal of carbonates; (iii) concentrations of sesquioxides; (iv) alterations that form silicate clay; (v) formation of granular, blocky, or prismatic structure; or (vi) combinations of these. In soils without organic surface accumulations, microbial activity tends to be maximized in the A horizon and declines precipitously in the B horizon.
1.1.2 Soil Formation and the Microbial Community
Soil microbiologists frequently report results of study of a limited number ofsoil types and provide minimal descriptions of the sites from which the soil samples were collected. It is not unusual to see statements such as “a garden soil was used” in the literature. From the foregoing discussion, it is apparent that an understanding of not only the currently existing physical and chemical properties of the soil but human‐imposed as well as natural events influencing soil formation must be examined. Furthermore, primary goals of research include not only improvement of our understanding of the function of the local soil site but also elucidation of the soil science principles that will increase our understanding of soils in general. Indicating the ecosystem type from which a particular soil sample was collected is generally inadequate to allow proper interpretation of the data, extrapolation of the principles revealed by the research, and, frequently, replication of the study by other researchers asking similar scientific questions. Although it may seem adequate to report that a grassland soil was evaluated in a particular experiment, application of proper soil classification procedures may reveal that a variety of soil types occur within the region classified by an ecologist as a grassland. That is, to conduct a meaningful examination of a soil ecosystem, the area to be sampled must be appropriately described. An appreciation of the variation of soil properties and the importance of the variation to the ecosystem services provided by the soil microbes can be gained by a short consideration of the controllers of soil genesis.
State Factor Theory of soil development: The effects of interactions of total ecosystem properties on soil development were described succinctly by Jenny (1941) in his State Factor Theory, frequently referred to as the CLORPT model. The original model stated that soil development is a function of climate, organisms, relief (topography), parent material, and time. The theory was subsequently modified to include human influences (Amundson and Jenny 1991). Although the model emphasizes the major factors affecting soil formation, possibly primarily from a macro‐environment view, it is reasonably clear that each of the factors affects and is affected by the soil microbial community. The synergistic associations of soil microbes and their physical and chemical environment determine the properties of all soils involved as well as the ecosystem services derived from the soil. The effect of some of the factors, such as climate, on soil microbes is clear, especially when comparing such extremes as desert and forest ecosystems or even boreal and tropical rainforests, whereas others such as relief are less obvious. Perhaps the latter effects on microbial function could be clarified by consideration of variation of temperature and moisture on east versus west aspects of a hill, where soil temperature would vary due to different impacts of morning and afternoon sun exposure or direction of prevailing rain storms.
In regard to human impacts on soil formation, at one point in human history agriculture could have been considered to be the primary controller of soil development (e.g. destruction of soil structure, alteration of nature and properties of organic matter inputs) but now it is reasonably easy to accept the observation that the genesis of all soils is directly affected by human society (e.g. distribution of pollutants, variation in air and water contents, as well as alteration of climate via anthropogenic‐imposed climate change or even drainage of large swamps). In all cases, the dynamics and nature of the soil microbial community are determined at least in part by the evolving soil properties as much as the evolution of the soil properties are determined by the function of the soil microbial community.
Interrelationship of microbial communities throughout the soil profile: A complication to description of soil systems results from the fact that soils are continually changing. To the casual observer, it may seem that soil types and their properties represent a constant in their conceptualization of an ecosystem. In reality, as can be concluded from the CLORPT model, soil physical, chemical, and biological properties are continually changing. Soil pH, salinity, aggregate structure, and cation exchange capacity are but a few examples of soil properties constantly evolving due to the actions of the biological community (plant, animal, and microbes). For example, microbes produce organic acids, carbon dioxide, and hydrogen ions that alter the soil pH, thereby affecting the solubilization of soil minerals and ultimately the salinity of soil water. Additionally, as a result of variation in the composition and production of root exudates, aboveground differences in plant community type and density can cause variations in belowground microbial and enzymological activities that may range over several orders of magnitude. This effect of plant biomass productivity on soil enzymes is exemplified through analysis of enzyme activity in a reasonably uniform organic soil (i.e. a single soil type) in the Everglades Agricultural Area (South Florida, USA). Acid phosphatase, invertase, xylanase, cellulase, and amylase activities varied by as much as 50‐fold between not‐cultivated and soils cultivated to sugarcane, St Augustinegrass (Stenatophrum secundatum [Walt] Ktze.), or paragrass (Brachiaria mutica [Forsk.] Stapf) (Duxbury and Tate 1981). A limited understanding of the complexities of these organic soil‐based systems would result if each soil sample (uncultivated or from grass or sugarcane fields) were simply classified as agricultural soil. Inputs from the aboveground plant community necessitate dividing the study area into at least three ecosystem types. Note also that in the study cited, all aboveground biomass could have been grouped into a category of grasslands (i.e. both sugarcane and the St Augustinegrass are by definition grasses).
Although plant communities are instrumental in soil development, it must be stressed that soil properties can be very different in seemingly related ecosystem types. This situation is exemplified by comparing two large forest groupings: temperate and tropical forest soils. Temperate forest soils may be moderately acidic (pH 3.5–4.5) and possess a well‐defined horizonal development (Figure 1.4). They may also have a surface litter layer plus a subsurface horizon (spodic horizon) with an accumulation of organic matter. Clearing of forests and cultivation of such soils in the temperature regions has resulted in moderately productive agricultural systems. In contrast, tropical rainforest soils generally lack a surface organic horizon; are usually highly acidic, nutrient‐poor soils; and can be poor candidates for agricultural development.
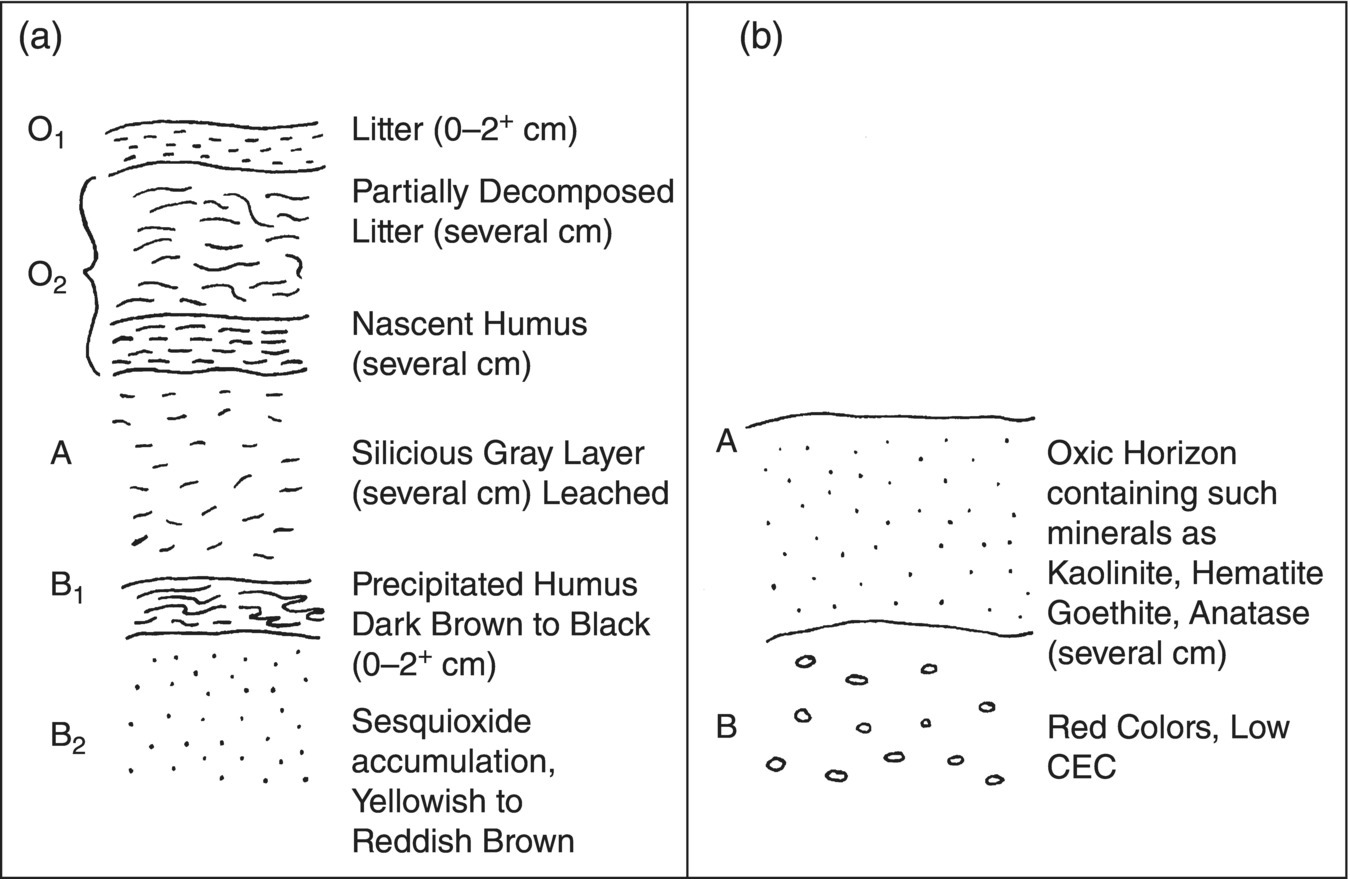
Figure 1.4 Comparison of idealized soil profile models for temperate and tropical forest soils. (a) Spodosol surface horizons with accumulated organic matter, and leached A horizon. Spodosols are exemplified by acidic forest soils. (b) Latosilization as would be seen in an oxidol (tropical forest ecosystem). These soils are generally highly leached, acidic soils with no significant humus layer. See Fanning and Fanning (1989) for further discussion of these soil types.
Thus, it may be concluded that our operational definition of a soils must include the concept that they are composed of unconsolidated mineral or organic matter, as stated above, but it must also be understood that general ecosystem properties affect soil property evolution. The microbiologist must realize that soil is a continuously developing entity with properties reflective of the site in which it has evolved and is still evolving. A corollary to this maxim is that significant changes in the overt soil properties will meaningfully affect the nature and properties of the soil microbial community. Ultimately, when characterizing a soil ecosystem, the history of the soil must be considered along with the current situation.
1.1.3 Implications of Definition of the Soil Ecosystem
Generally, understanding the effect of microbial/soil mineral interactions is important for developing local soil management plans, but a more general goal of studying soil processes could be to elucidate the underlying principles of soil development, enabling extrapolation of the data to larger soil systems or even soils in general. The ability to extrapolate experimental findings to a larger venue depends on the extent of the soil system selected for study. Frequently, the greater the variability of physical or chemical properties in the soils and the better the principles underlying their impact on biological properties are elucidated, the greater the applicability of the discoveries to our understanding of the properties of soils in general. Unfortunately, simply because of the logistics involved in soil sampling and laboratory analyses, arbitrary limitations of the extent of the ecosystem area studied are inevitable.
Variability of soil properties within even a more limited field site is such that a basic requirement of research projects is that the soil samples must reflect site variation. The better our understanding of soil variability, the better the potential for elucidating the full range of the microbial processes occurring therein and the properties of the soil controlling these activities. Conversely, the smaller the portion of the overall system studied, the less the potential for valid extrapolation of the data to provide a description of the total ecosystem. Thus, a soil microbiologist faces the dilemma of selecting a soil site small enough to be sampled in a representative manner without being overwhelmed by the number of soil samples to be studied, but sufficiently expansive that the data collected will be at least to some degree representative of the total ecosystem. The more limited the dimensions used to define an ecosystem of consideration, the more important becomes consideration of the impact of external processes on the integrity of the system. For example, a B horizon soil may be examined, but it must be considered that energy derived from oxidation of organic soil components depends to a large degree on the rate that the fixed carbon is leached from the A horizon to the B horizon. Drawing boundaries that define the portion of the larger ecosystem to be studied will clearly enable some aspects of the ecosystem to be self‐contained (that is, be treated as a closed system) whereas others must be supplied from external sources (that is, function as an open system). A good example of this situation involves the aerobic microbial populations deriving their carbon and energy from oxidation of organic carbon (the heterotrophs). In the vast majority of situations, the carbon oxidized for energy originates from beyond the world of the bacterial micro‐colony. It is produced photosynthetically by aboveground plant communities.
1.2 The Micro‐ecosystem
Existence of aboveground communities literally rests on the stability and function of soil processes occurring at the microscopic level. Because microorganisms range in size from less than a micron to a few microns, individual soil particles have a major impact on their function. The challenging aspect of this observation is that the biological processes occurring at the micron level can have profound implications on total ecosystem development and longevity. For example, a series of bacterial colonies consisting of as few as a half‐dozen cells each may determine the fate of a potentially ecosystem‐destructive xenobiotic compound or the bioavailability of toxic metals, such as mercury or arsenic. Thus, a clear understanding of the interactions of the soil microbial community and elemental soil components (sand, silt, clay, and colloidal organic matter) is imperative to comprehend total ecosystem function.
1.2.1 Interaction of Individual Soil Components with the Biotic System
Surface area and charge density have major effects on microbial and enzyme activity in soil. The basic components of soil are sand, silt, clay, and humic substances. Although all soil particulates provide surfaces for microbial colonial development, clay and colloidal organic matter have the smallest diameters and therefore present the largest surface area for interaction with soil microbes and their products. The relationship between estimated particle diameter and surface area is demonstrated in Figure 1.5. Due to their colloid size and the fact that humic acids and most clays are general negatively charged, these two soil physical components are major determinants of the extent of activity of soil microbes, enzyme activity and availability of their substrates. Thus, the nature and properties of these soil components must be examined in greater detail.
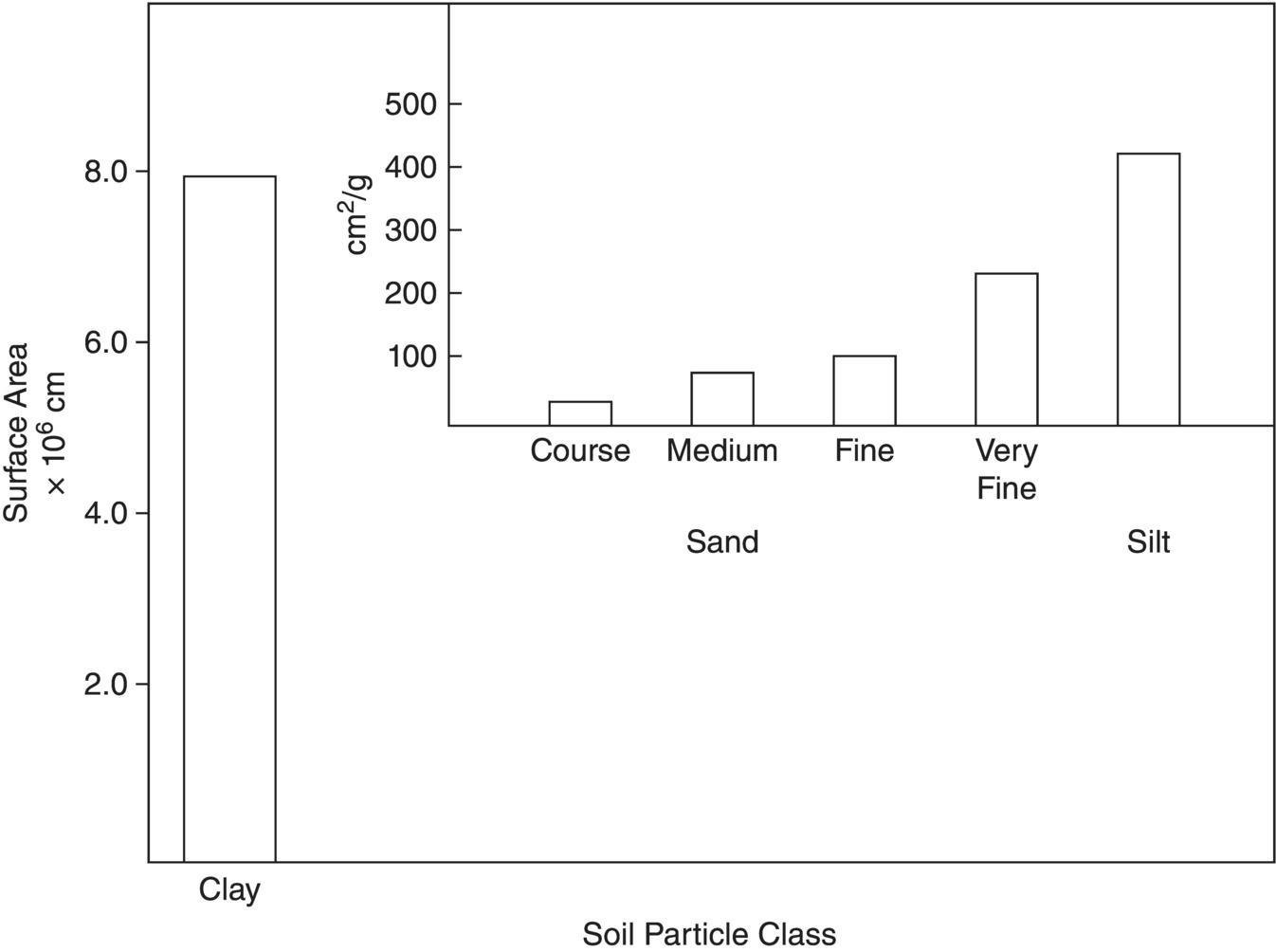
Figure 1.5 Variation of surface area of soil particles: The particles are assumed to have spherical shapes.
Source: Data are selected from Foth and Turk (1972).
1.2.1.1 Clay and Ecosystem Function
The high negative charge and high surface area g−1 clay result in the clay particles having a unique impact on the soil biological and chemical processes. The negative charge of 1 g of clay particles is at least 1000‐fold that of 1 g of sand (Figure 1.5). Thus, even if the clay particles constitute as little as one‐tenth of the mass of soil particulate matter, they have a controlling role in soil biological, chemical, and physical processes. Soil water relationships provide an excellent example of the nature of these clay‐driven interactions. Soil microbes are commonly associated with the surface of soil particles. Water films are distributed around the soil particles. Due to the physical interactions between the bipolar water molecules and the soil particle surfaces, water molecules closest to the clay particle surface are held more tightly to the clay particles than they are to each other in the water layer, so are less available to the microbes (see Brady and Weil (2017) for further description of the role of hydrogen bonds and dipole moment in these processes.) This negative charge is sufficient to create water – clay layering in the area adjacent to the clay particles. Water – water layering occurs further out from the particle surface to result in a clay – water + clay − water layering effect. In soils with more surface areas and high negative charges, more water is needed to coat the particles before a sufficiently thick water layer on the clay particles for the microbes and growing plants is formed. Again, refer to Brady and Weil (2017) for further information regarding these processes and interactions.
Considering the disproportionate impact on soil processes that results from the colloidal size/high surface area of clay minerals, it can be concluded that an examination of basic clay properties is key to understanding microbial function in soils. Clay minerals are secondary minerals; that is, except for mica, clay minerals are formed in soil from the weathering of primary minerals or from the products of their weathering. Most clays are crystalline and are composed of layers of oxygen atoms associated with silicon and aluminum atoms through ionic bonding. For crystalline clays, these silicate and aluminum layers are stacked (like a deck of cards) into clay particles, termed micelles. As will be noted below, less regularly oriented amorphous clays are also found in some soils. Common groups of clays are layer silicate clays, hydrous oxides of iron and aluminum, plus allophane and other amorphous clays.
Layer silicate clays are primarily crystalline clays with specific structures and predictable properties. These clays have a plate‐like structure formed from flat sheets of silicon, aluminum, magnesium, and iron linked by oxygen and hydroxyl groups. The major silicate clays are commonly referred to as phylosilicates due to their leaf‐like structure. The crystalline structure can be summarized as being composed of two distinct crystalline sheets.
· A silica‐dominated sheet composed of silicon atoms surrounded by four oxygen atoms in a tetrahedryl arrangement. The interlocking array of silicon and shared oxygen atoms is a tetrahedron sheet.
· An aluminum‐ or magnesium‐dominated sheet composed of aluminum and magnesium atoms surrounded by six oxygen atoms in an octahedryl arrangement. Thus, the layer is referred to as an octahedryl sheet. If the layer is dominated by aluminum, it is a dioctahedryl sheet, and if by magnesium, a trioctahedryl sheet.
The tetrahedryl and octahedryl sheets are bound together in regular crystals by shared oxygen in the different layers. Major groups of these layer silicate clays are:
· 1 : 1 – one tetrahedryl (Si) sheet per octahedryl (Al) sheet. Kaolinite is the most prominent member of this group
· 2 : 1 – one octahedryl sheet sandwiched between two tetrahedryl sheets as exemplified by smectite (montmorillonite)
· 2 : 1 : 1 – typically 2 : 1 crystals as above alternating with Mg‐dominated trioctahedryl layers as exemplified by chlorites
Both a variable (depending on soil pH) and a fixed negative charge are associated with layer silicate clays. The pH‐dependent charge results from ionization of the hydroxyl groups on the surface and edges of the crystal. The fixed charge is the product of isomorphous substitution for the aluminum or silicon atoms within the crystalline structure. For example, the aluminum atom is only slightly larger than the silicon atom. Therefore, aluminum can fit into the center of the tetrahedron in place of the silicon without altering the basic physical structure of the crystal. The aluminum atom carries a +3 charge compared to the +4 charge of the silicon. Thus, one of the negative charges of the shared oxygen atoms is not satisfied, resulting in a net negative charge of the crystal.
The negative charges of clay minerals contribute to the soil cation exchange capacity and are instrumental in interactions with soil organic components and microbes. The quantity of negative charges associated with clay minerals is a property of the specific clay type. Thus, identification of the type of clay in a soil site is essential for predicting the chemical and biological dynamics occurring therein.
Gibbsite (Al203.3H20) and goethite (Fe203.H20) are examples of hydrous oxides of iron and aluminum. These clays are found primarily in the highly weathered soils of the topics and subtropics, but also occur in temperate regions. They are less studied than the layer silicate clays. Hydrous oxides of iron and aluminum may be crystalline or amorphous. They are not sticky clays and their charge varies with pH. At high pH, they have a slightly negative charge, whereas at the highly acidic pH values common to the weathered tropical and subtropical soils where they generally occur, they are positively charged. This contrasts to the layer silicate clays, which are normally negatively charged.
Allophane and other amorphous material clays are much less studied than the two other clay mineral groups. The most significant members of this class of clays are allophane and imogolite. They are aluminum silicates and are prevalent in andisols (volcanic ash‐derived soils). Thus, examples of locations where they could be encountered are the Northwestern United States, Hawaii, and Japan.
Adsorption of organic substances to clay particles: Clay minerals provide a surface for sorption of a variety of organic substances. Not only do whole microbial cells and viruses sorb to soil clay particles, but also a diversity of organic compounds have been demonstrated to interact with clay surfaces. Examples of the types of organic compounds reacting ionically with clay particles include carbohydrate phosphates (Goring and Bartholomew 1951), nucleotides (Goring and Bartholomew 1952), humic and fulvic acids (Greenland 1971; Kodema and Schnitzer 1974) and aromatic compounds as exemplified by p‐cresol (Boyd and King 1984). Most biologically produced compounds interact with clay particles. The environmental quality concerns relating to clay–organic matter interactions involve the bioavailability of the sorbed substances and, for biodecomposable substances, the impact of sorption on degradation kinetics. Tightly sorbed organic substances, especially those existing within soil micropores, may be essentially unavailable to the biological community, i.e. they may be sequestered. Biodegradation susceptibility of sorbed organic compounds may be reduced or enhanced. Thus, the adsorption potential of any organic compound entering soil should be evaluated in relation to estimation of the effect of this process on its longevity in situ.
It must be emphasized that both substances entering the soil system from external sources (e.g. pesticides and other soil amendments) and those synthesized in situ may be sorbed onto clay surfaces. Sorensen (1972) found that amino acid metabolites synthesized in montmorillonite‐amended soil following cellulose, hemicellulose, or glucose amendment were bound by soil clays. Clay‐amended soil retained two–threefold more carbohydrate‐derived amino acid carbon than did nonamended soil over a three‐year incubation period. In a related study, 14C‐labeled cellulose was added to seven soils with silt plus clay contents ranging from 8% to 75% (Sorensen 1975). As in the previously cited study, the quantities of amino acid carbon synthesized from the cellulose remaining in the soil after 30 days of incubation ranged from 6% in the lowest silt plus clay‐containing soil to 18% in the highest silt plus clay‐containing soil, implying a stabilizing or protective effect due to sorption to clay minerals. This stabilizing effect by clay minerals on organic matter is of particular interest when considering the potential for mitigating the climate change potential resulting from mineralization of soil organic matter.
Association of organic compounds with clay particles is not a totally random process. There is an enrichment of specific carbon moieties within clay particles. This is exemplified by evaluation of the properties of humic acids associated with clay particles. In native soil samples, aliphatic structures are more common in humic acids derived from the clay fraction than other soil fractions (Arshad and Lowe 1966; Schnitzer et al. 1988; Schulten and Schnitzer 1990). Using chemical, infrared, and 13C nuclear magnetic resonance (NMR), and gas chromatographic mass spectrometric analysis, Schnitzer et al. (1988) found that about half of the clay‐associated organic matter consisted of humic materials, the remainder being mainly long‐chain aliphatics.
Clay–organic matter adsorption mechanisms: An immediate conceptual problem raised by this list of materials sorbed to clay particles is the observation that both the most commonly occurring clays in temperate region soils and many of the sorbed organic compounds are negatively charged at the predominant soil pH. Thus, mechanisms must exist to reduce the repulsive forces between the like charged entities. Examples of such mechanisms are reduction of the quantity of negative charges (pH modification or methylation of the carboxyl groups on the organic matter) and cationic bridging of negatively charged functional groups of the humic substances with the clays. The latter process is depicted by a model proposed by Greenland (1971) (Figure 1.6). The negative charges on the organic and mineral materials are linked in soils by divalent cations.
Organic compounds retained on clay particles through cationic bridges can clearly desorb from the surface with changes in ionic strength and cation content of the soil solution. A more permanent association of organic compounds to clay particles can occur. This association between clay particles and organic materials results from interlayer clay–organic matter complex formation. For example, this was demonstrated to occur in two New Zealand soils by Theng et al. (1986). Humic substances with a polymethylene chain structure were found to be regularly interstratified with mica‐smectite in highly acidic soils containing low microbial activity.
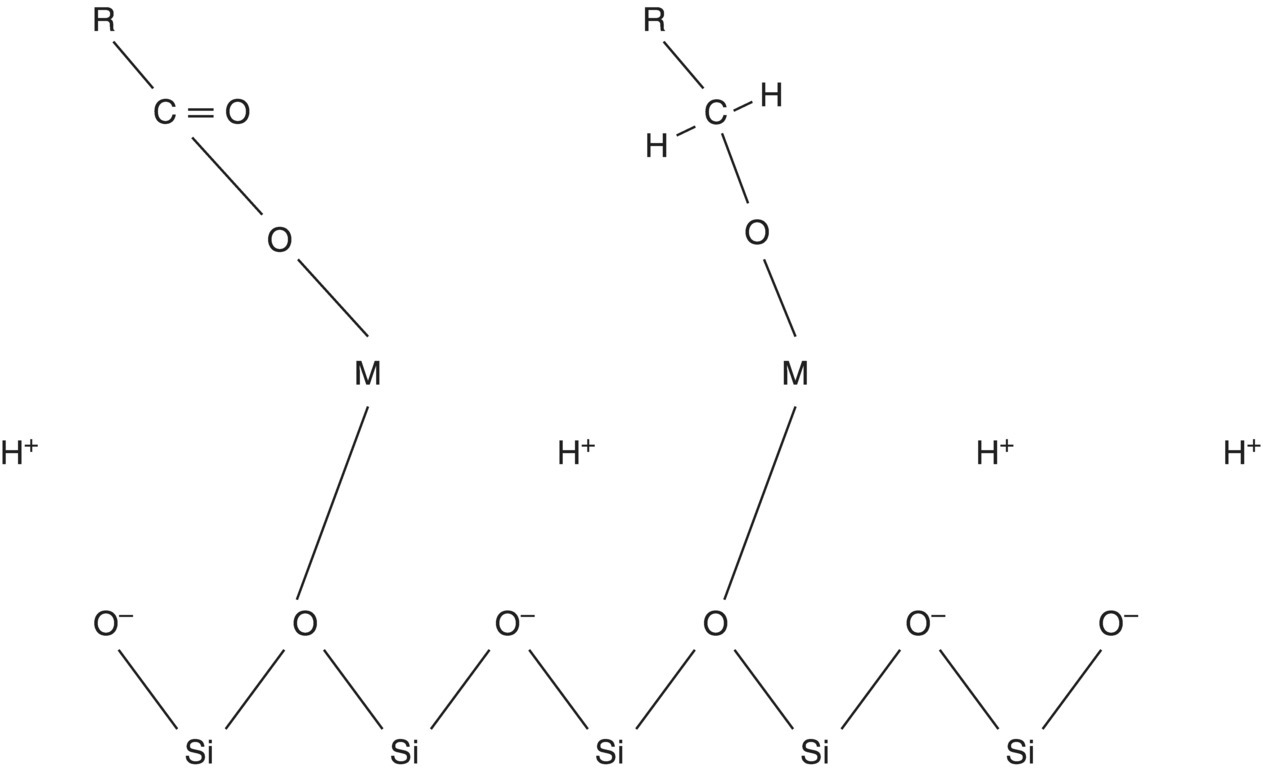
Figure 1.6 Schematic drawing of divalent metal (M) salt bridge linking of the surface of a mica type clay with a negatively charged organic compound and the distribution of a hydrogen ion cloud around the clay surface.
Any process that alters the negative charges of either the clay or organic matter reduces the repulsive forces. This can be accomplished by altering soil pH or through methylation of the carboxyl groups. For example, Nayak et al. (1990) demonstrated that reduction of the acidic groups of humic substances through methylation increased fixation with montmorillonite. The increased association with the clay particles resulted not only from a reduction of interparticle repulsive forces but also from a reduction in hydration energies.
Biodegradation and clay interactions: It has long been understood that the association of clay and soil organic matter affects the decomposition rate of sorbed organic compounds. For example, early studies of the impact of clay on decomposition of a wide variety of organic compounds have shown that purified clay minerals when added to growing microbial cultures can stimulate, decrease, or have no effect on decomposition of organic materials (see Hwang and Tate 1997; Kunc and Stotzky 1974). Kunc and Stotzky (1974) found no clear relationship between the type of organic substance and the type of clay relating to the effect on biodegradation except with aldehydes, whose decomposition was accelerated by clay minerals. Lynch and Cotnoir (1956) noted that bentonite inhibited decomposition of a variety of substrates but that illite and kaolinite had little effect on decomposition. Similarly, Olnes and Clapp (1972) found that the kinetics of the decomposition of the montmorillonite‐dextran complex contained periods that were affected to a greater degree by the montmorillonite content, whereas other portions of the degradation curve were affected more by the dextran concentration.
Clearly, it is difficult to make general statements regarding the impact of clay sorption on biological decomposition kinetics. The basis for these seemingly contradictory observations can be revealed by examining the effect of clay on bioavailability of sorbed organic compounds. Two ramifications of clay–organic matter interaction can be pictured depending upon the relationship of the quantity of substrate present in the system and the ability of the organism to interact with it. Carbon and energy sources for microbes may exist in their ecosystem at concentrations too low for significant interaction between them and the microbes to occur. These materials may become sufficiently concentrated on the clay surface that the quantities desorbed from the clay and thus occurring in the water surrounding the clay particle are sufficient to induce microbial catabolism. In contrast, for substrates present in high concentration in soils with high clay contents, their concentration in solution could be reduced by adsorption to the clay particle to an extent sufficient to preclude or greatly reduce the rate of decomposition. In this latter situation, strong binding of the carbonaceous substance to the clay particle reduces the probability of its collision with and uptake by the degrader population. The rate of desorption from the clay thus becomes the limiting factor determining the rate of decomposition of the carbonaceous energy source. Thus seemingly contradictory observations as suggested above could be reasonably explained by examining the chemical processes associated in the partitioning of the carbon and energy source between the bound and soluble nutrient pools and the effect of proportion of the organic substances in the soil that is bioavailable.
Thus, the impact of clay sorption in soil ecosystems is more a function of residual concentrations of organic compound in interstitial waters and in the microsite containing the microbial colony than is the apparent concentration when the bulk soil sample is analyzed (i.e. microsite concentrations of the compound of interest may far exceed macrosite levels). Thus, decomposition kinetics of the substance reflect the microsite concentrations rather than those that would be predicted from total ecosystem content of the carbonaceous growth substance.
Not only do clay minerals alter decomposition kinetics of organic substances in soil through sorptive‐desorption reactions, but they may also indirectly affect microbial metabolism by altering their chemical environment. For example, due to the formation of an hydrogen ion cloud around the clay particle, the pH of the microsite is reduced (see Figure 1.7). Thus, enzyme‐catalyzed reactions in clay soils may appear to have pH optima lower than that detected in the absence of clay particles.
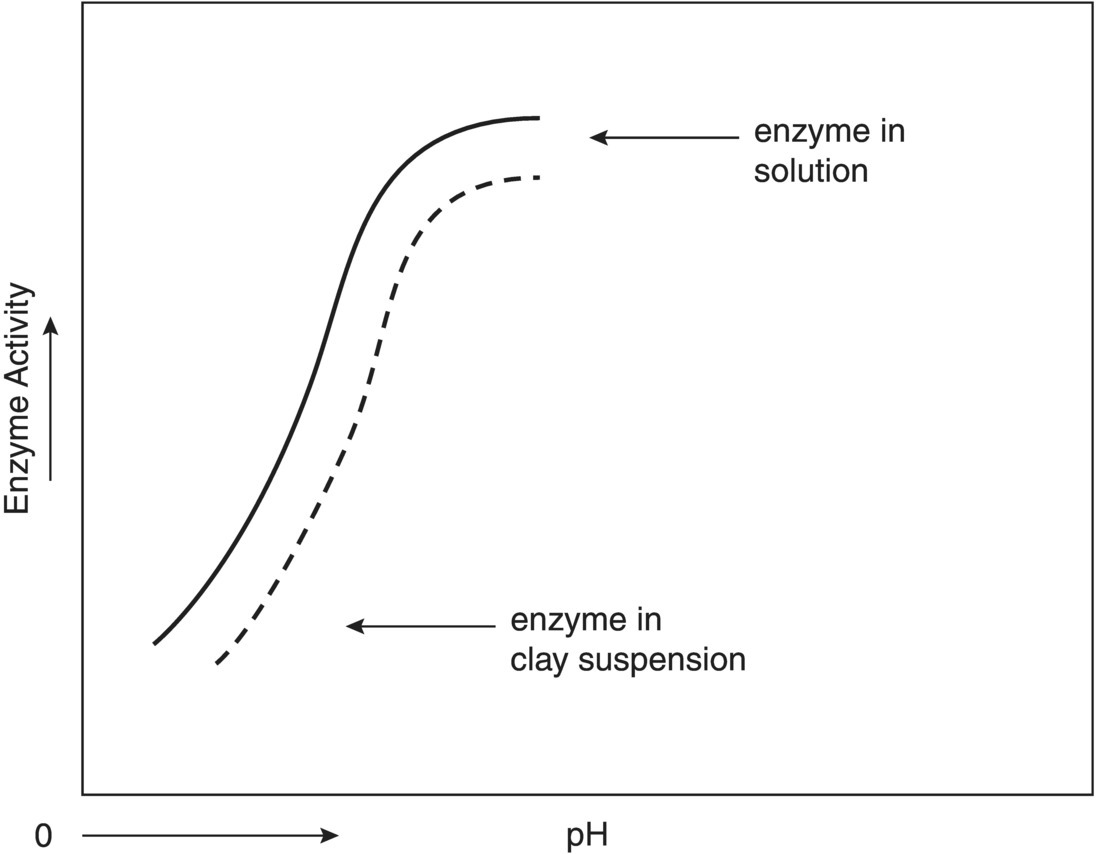
Figure 1.7 Hypothetical data reflecting alteration of apparent pH optima by microsite variation in pH due to clay micelles.
1.2.1.2 Humic Substances and Ecosystem Function
Soil organic matter is commonly considered to be equivalent to soil humus (Stevenson 1994). The soil humus fraction consists of all organic compounds normally found in soil (with the exception of recognizable plant or animal components), their partial decomposition products, and microbial biomass. Studies of the nature of soil organic matter are complicated by the diversity of materials present and the fact that the definition does not coincide with what can be reasonably separated and analyzed; for example, clear separation of plant and animal biochemicals from the bulk soil organic matter is not easily accomplished. The mixture of chemical substituents of soil humus ranges from simple monomeric components of living cells, such as organic acids, amino acids, and saccharides, to the random polymers classified as humic acids. Essentially any organic compound contained in or synthesized by living cells occurs in soil, although some may be present in minuscule amounts due to their being relatively easily decomposed by soil microbes.
Soil humic substances: The conclusion that soil humic substances have a major input in determining the fundamental soil properties demands a deeper consideration of the basic properties of these substances. By definition, humic substances are a series of high molecular‐weight, brown to black substances formed by secondary synthesis reactions in soil. This organic matter fraction is not designated or isolated from soil based on a function or role in the soil ecosystem. It is a portion of soil humus that is defined strictly by its solubility in acidic or alkaline solutions. Humic substances are separated from soil with alkaline solutions. Acidification of the alkaline extract to a pH between 1 and 2 results in precipitation of the humic acids. Fulvic acids are soluble in both acid and alkali. Humin is that portion of the colloidal soil organic matter fraction that is not solubilized with alkaline solutions
To understand the impact of humic substances, especially humic acids, on microbial function in the soil ecosystem, an appreciation of the structural components interacting with soil particles and the microbial community is necessary. Humic and fulvic acids are heterogeneous, random chemical associations of a variety of aromatic and aliphatic components. No single chemical structure can be proposed to represent these molecules. Indeed, because a wide diversity of aromatic and aliphatic compounds are linked via a variety of covalent linkages forming humic acids with molecular weights in the thousands of daltons, it can reasonably be concluded that few if any of the molecules are identical. In contrast, fulvic acids are more oxidized than humic acids; that is, they have higher oxygen content and a lower carbon content and molecular weight.
Structural implications of humic acids: The nature of the constituents of humic substances determines their importance to total ecosystem function. The heterogeneity and complexity of humic and fulvic acids have limited the level of understanding of the specific chemical reactions between humic and nonhumic soil organic components. An appreciation of the types of reactions occurring between microbial cells or individual biochemicals and humic acids has developed from our basic concept of the molecular content of humic substances and the use of related model compounds.
Our concept of humic acids structure and in situ interactions in soil was improved through the application of spectroscopic analytical techniques, such as NMR spectroscopy. (See Preston (1996) for a review of the utility and limitations of NMR techniques in elucidating the elements of humic acid structure.) Data derived from NMR‐based analytical procedures suggest that aromatic rings make up a smaller portion of the humic acid molecule than was previously believed (e.g. see Hatcher et al. 1981; Malcolm 1990). Hatcher et al. (1981) found that aromaticities of humic acids from a variety of soils collected from Prince Edward Island ranged from 35% to 92%. Significant aliphatic structures were generally noted. These were not detected by chemical oxidation procedures. These authors also suggested that phenolic carbons may be minor components of humic acids. They were previously considered to be major substituents. This conclusion was based on the observation that NMR spectra did not contain intense signals corresponding to the presence of these molecules. Schnitzer and Preston (1986) suggest that since the presence of phenolic hydroxyls is confirmed by several independent methods, failure to detect them by NMR analysis merely indicates a shift in the NMR signal resulting from these moieties.
With the development of more sensitive methods for analysis of humic acid structure, a clearer understanding of the interactions of these substances with soil minerals as well as xenobiotic chemicals is becoming possible (for more information on this topic refer to Cao et al. 2011; Simpson 2006). Particularly of interest in determining the capability of xenobiotic chemicals to be sequestered within humic acids is the debate regarding the total size of the humic acid molecules and whether they are composed of aggregates of smaller molecules, single very large molecular weight molecules, or a combination of both (e.g. see Cao et al. 2011; Li et al. 2003; Piccolo et al. 2002).
The primary conclusion regarding the structure of humic substances and their importance to the soil microbiologist is derived from the complexity of their composition. They contain varying ratios of phenolic, methoxyl, aromatic, hydrocarbon, amino acid, and nitrogen moieties plus covalently linked polysaccharides and proteins. The ratio of these substituents is dependent on the source of the humic acid. Great variation in composition occurs between humic acids prepared from soil, stream, and marine samples (Malcolm 1990). Significant differences in humic acids have also been noted for humic acids prepared from a variety of soil types – alfisols and mollisols (Novak and Smeck 1991) – and soil management histories (Stearman et al. 1989).
Humic substances, organic compounds, and biodecomposition: Although a variety of microorganisms have been suggested to be capable of metabolizing humic acids, it is unlikely that humic acids are meaningful contributors to the energy resources of soil microbes. This conclusion results primarily from observations of the complexity of the structure of molecules – they contain a large variety of substituents that are essentially randomly assembled. Therefore, an inordinate amount of metabolic energy would have to be expended in synthesizing the large array of degradative enzymes needed to decompose the humic acids – considerably more energy than the microbe would recover from mineralization of the complex molecule. Humic substances affect soil biological processes to the greatest extent by interacting with soil minerals in soil aggregate formation (see Section 1.2.2) and by their role in removal of biodegradable substances from the pool of easily metabolized substrates (i.e. humification). Enzymatic interaction with humified substrates is inhibited physically by the imposing humic acid physical structure.
As was introduced above in the discussion of clay/organic substance interactions, humic substances sorb organic compounds through temporary interactions. For these organic matter interactions, sorption results from n–n interactions, van der Waals forces, and hydrogen bonding as well as ionic associations. More permanent linkage of organic compounds to humic substances also occurs due to formation of covalent bonds. Examples of organic compounds that may be linked covalently to humic acids include amino acids, peptides, proteins, aromatic compounds, and polysaccharides.
Covalent linkage of phenolic compounds to humic acids may be catalyzed by phenoloxidase (Bollag et al. 1980; Dec and Bollag 1997; Liu et al. 1985). Carbon–carbon and carbon–nitrogen linkages are catalyzed through free radical formation by these laccase‐type enzymes (Figure 1.8). The linkage of aromatic ring‐containing compounds occurs in ortho‐ortho or para–para or ortho‐para type associations by this biologically catalyzed process (Sjoblad and Bollag 1981). Aside from being a mechanism for the formation of humic substances in situ, these oxidative processes provide a means for the stabilization of a variety of xenobiotic compounds, such as pesticides, into soil organic matter.
The impact of the covalent association of organic compounds with humic substances on their biodecomposition kinetics is more predictable than was described above for clay–organic matter interactions. Humification of organic compounds results in their stabilization in soil. For example, in a study of the impact on microbial degradation by nucleophilic addition of amino groups to the aromatic nuclei, Bondietti et al. (1972) found that bonding of amino sugar units to model humic acids resulted in major reductions in the decomposition rate. Over a 12‐week incubation period, more than 70% of free glucosamine was oxidized to carbon dioxide, while only 15–23% of the humic acid‐model polymer‐bound amino sugar was oxidized. The importance of covalent linkage of the humic acids and sugar was shown by the fact that simply mixing the amino sugar with the model humic acid molecule did not reduce the oxidation rate.
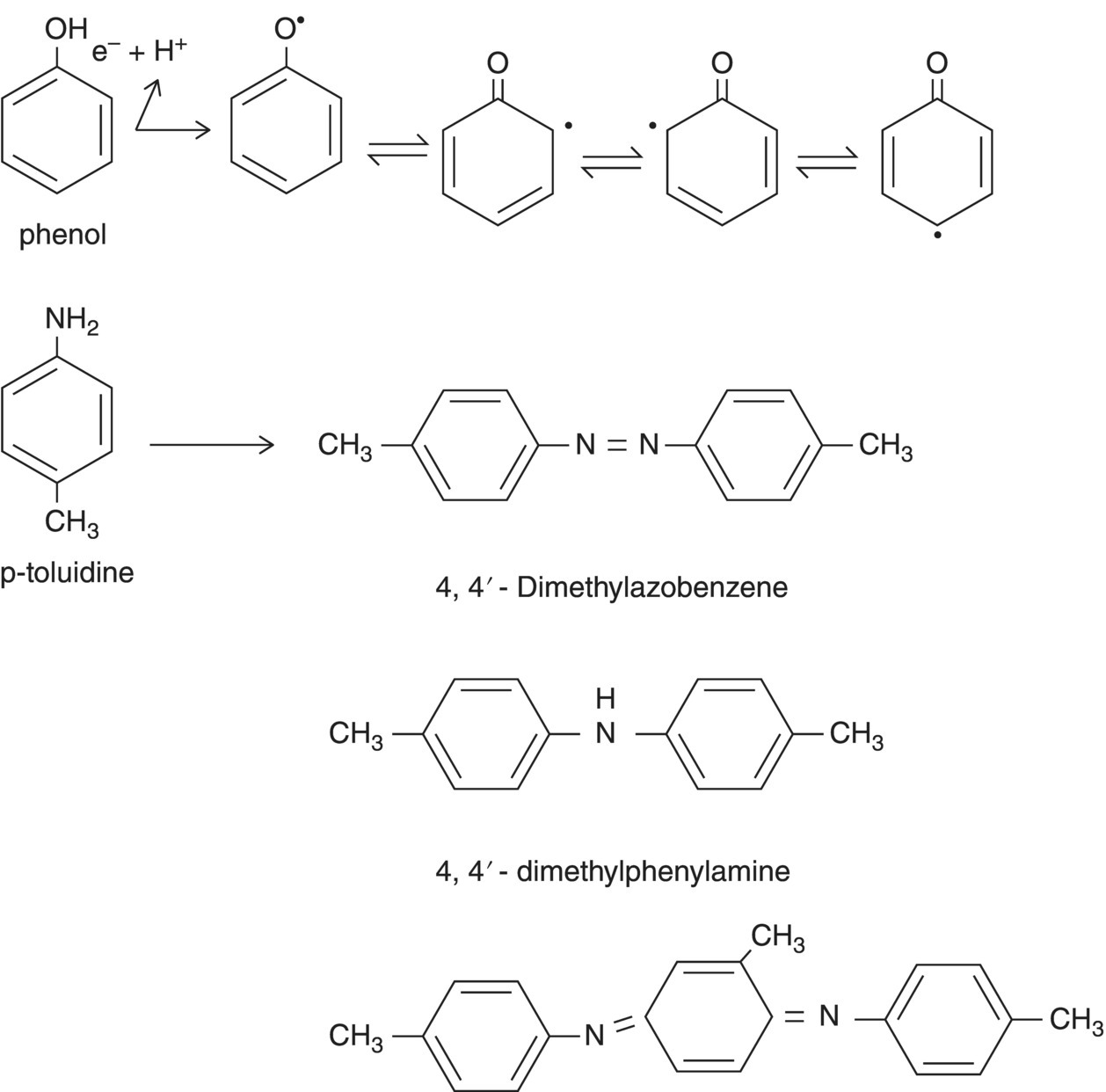
Figure 1.8 Oxidative coupling of aromatic compounds. Mechanism showing formation of free radicals and examples of some products of p‐toluidine polymerization. See Sjoblad and Bollag (1981) for further discussion and examples.
It is important to note that substances covalently bonded to humic acid can still be mineralized but that the rate of oxidation is much slower than for nonchemically linked substances. Basically, what is under consideration here is the fate of the carbon of the humified organic compound after it has been incorporated into the humic acid molecule. Since it is part of the larger substance – that is, humic acid – its decomposition kinetics tend to resemble the slow mineralization rate of humic acid molecules. This observation has environmental significance. Covalent bonding to humic acids could be a viable mechanism for renovation of xenobiotic compound‐contaminated soils if the substances are permanently bound or if they are released in an inactivated (i.e. partially or totally catabolized) form. Unfortunately, this condition is not always met. Release of humic acid‐bound pesticides intact can occur (Bartha 1980; Lichtenstein 1980; Still et al. 1980). Thus, from an agricultural viewpoint, difficulties may arise from release of a pesticide that had been previously applied to an appropriate crop and stabilized in soil humic substances several years after application. Particular concern would arise should this pesticide reappear in a crop where use would be inappropriate from either a crop sensitivity or regulatory view. Additionally, incorporation of photosynthetically fixed carbon into humic substances and associated molecules mitigates at least a portion of the impact of carbon dioxide production on global climate change.
It must be stressed that in most cases, the more probable fate of a biodecomposable substance added to soil is mineralization, either before humification or even subsequent to humification. For example, Haider and Martin (1988) found that Phanerochaete chsysosporium could mineralize xenobiotic compounds at a reasonable rate (conversion of 13–56% of xenobiotic carbon as carbon dioxide over an 18‐day period). Similarly, Saxena and Bartha (1983a) found that humus‐bound 3,4‐dichloroaniline (a biodegradation intermediate) was mineralized at a rate approximately the same as that of the humic acid molecule itself. Adsorbed 3,4‐dichloroaniline was mineralized faster than humic acid molecules. Of note is the observation that fertilization of agricultural soil with anhydrous ammonia could mobilize humus‐bound 3,4‐dichloroaniline (Saxena and Bartha 1983b).
The size and complexity of the humic acid molecule create the potential for a stable, noncovalent association (sequestration) between it and simple organic molecules. Using the classic formula for the humic acid molecule, Schulten and Schnitzer (1997) have described a three‐dimensional structure of the molecule that can accommodate noncovalent associations of simple organic molecules within voids in the molecule. These associations are sufficiently stable energetically that the longevity of the associations between the simple and complex organic substances resembles that of covalently associated molecules (Dec and Bollag 1997). Hydrophobic compounds have been shown to associate with hydrophobic domains, micelles, within the humic acid molecule (von Wandruszka 1998). Such associations may participate in reduction of bioavailability of toxicants through sequestration (see Chapter 16). Environmental concerns relating to these noncovalent associations relate to the need to assess the extent of their occurrence in native soil ecosystems, to evaluate the potential for release of the toxicant from the association, and to quantify the environmental risk resulting from the freeing of the toxicant from the humic acid mycelle.
1.2.2 Aboveground and Belowground Communities and Soil Ecosystem Synergistic Development
A primary controller of the structural associations of soil minerals is the interactions between soil microbes, higher plants, and soil particles. These interactions in turn control total ecosystem development, sustainability, and quality. A visual examination of a productive surface soil easily leads to the conclusion that the sand, silt, and clay components tend to be aggregated into larger associations – soil aggregates. Optimization of the soil aggregate structure produces a soil environment more supportive of plant growth. Thus, the question arises of how soil aggregates are formed.
A somewhat cursory consideration of the basic interactions between soil microbes, higher plants, and soil particles provides a foundation for understanding how these more productive structures come to be. The first consideration is that the soil microbial community derives the bulk of its cellular energy from the oxidation of fixed carbon produced via plant photosynthesis. Layered upon this fact is the observation that maximal plant biomass production relies in part on existence of a soil physical structure conducive for plant growth. The microbial community is a primary determinant controlling optimization of the soil structure for plant growth. For example, the soil structure must be such that water infiltration and oxygen diffusion are not impeded as is exemplified in the extreme situation by a compacted soil. In such a soil, soil quality and therefore plant biomass productivity are limited. This conclusion is predicated on the understanding that the evolution, growth, and sustainability of the plant community are determined by interactions of the soil microbial community with soil particulate matter. Soil bacteria and fungi are major players in the grouping of soil particulates into aggregates. A plant growing in a poorly aggregated soil provides only a limited amount of the carbon and energy required for sufficient microbial productivity to optimize soil aggregate structure. But, even in low productive ecosystems some carbon and energy are provided to the soil microbes by the limited plant growth; that is, sufficient energy to increase the level of formation soil aggregates at least minimally. The small improvement in soil structure could subsequently increase the amount of microbial biomass, so in turn if the soil is properly managed this results in improvement of the soil structure, which can allow for an increase in the ecosystem productivity. Two such systems – soil aggregate formation and biological soil crusts – exemplify how soil ecosystems are built upon these synergistic interactions.
1.2.2.1 Soil Aggregate Structure Development and Its Impact on Ecosystem Function
The world of the soil microbe does not consist of individual sand, silt, or clay particles or even of organic matter in toto. The microbial community must be capable of interacting with a mixture of these substances assembled in a seemingly random order. Yet when examined microscopically, the soil microorganisms may appear to be associated with one specific soil component in this complex environment. For example, Fisk et al. (1999), using a combination of specific stain and micromorphological techniques, found that soil‐amended bacteria tended to become associated with variably charged mineral oxides and organic matter within the intergrain microaggregates of a Freehold sandy loam and occurred along coated mineral surfaces. Also, rhizosphere organisms may be growing essentially on root surfaces. Microbes may be physically attached to partially decomposed plant debris. But even in these situations, the metabolism of soil microbes is affected by the association of the soil particle to which they are attached within larger soil aggregates. Note that the soil particles, the microbial community and the plant root system interact to provide a structure with enhanced water infiltration and retention, encourage the cycling of plant carbon, nitrogen, and phosphorus between belowground and aboveground communities, and facilitate optimization of a soil structure that supports ecosystem productivity and sustainability.
Soil structure is defined at the level of the microbial cell by the association of soil particulates and colloidal organic matter into soil aggregates. As noted previously, soil aggregates are combinations of soil organic and mineral components assembled into microaggregates (<50 μm mean diameter) and macroaggregates (>50 g mean diameter particles).
Soil aggregate formation: Assembly of soil particulates into aggregates is an excellent example of biotic and abiontic organic matter plus living cells and mineral components interacting to affect soil physical properties. The biological contributions to soil aggregate formation can be described as a coupling of feedback‐controlled processes (Figure 1.9). Biological participants are higher plants as well as soil microbes. The soil physical matrix is produced through the synergistic interactions of the biological entities associated with the soil particulates. Microorganisms metabolize photosynthetically fixed carbon to produce the compounds responsible for binding soil mineral matter together; that is, polysaccharides and humic substances. Humic acids are the product of aliphatic and aromatic compounds, some of which are biologically synthesized, being chemically and biologically linked into complex high molecular weight compounds. Improvement in the size and number of aggregates formed enhances plant community growth, which in turn stimulates even further microbial development by producing increased levels of fixed carbon. Soil quality and associated functions are enhanced in proportion to the degree of soil aggregation. Note that aboveground plant community development can be enhanced directly in proportion to the soil aggregate structure improvement depending upon the inherent production capacity of each species. In contrast, poor soil structure retards air diffusion, water movement and root growth, thereby resulting in a less active soil microbial community than would be attained under normal optimal growth conditions. Recall that mineralization of the plant biomass is the primary energy source for the soil microbes. Thus poorly aggregated soil would result in limiting fixed carbon inputs into the soil. As soil aggregate stricture is improved, the production of plant biomass allows for enhanced microbial biomass production. Additionally, optimization of the microbial decomposition of the fixed carbon improves the recycling of nutrients such as nitrogen back to the growing plants.
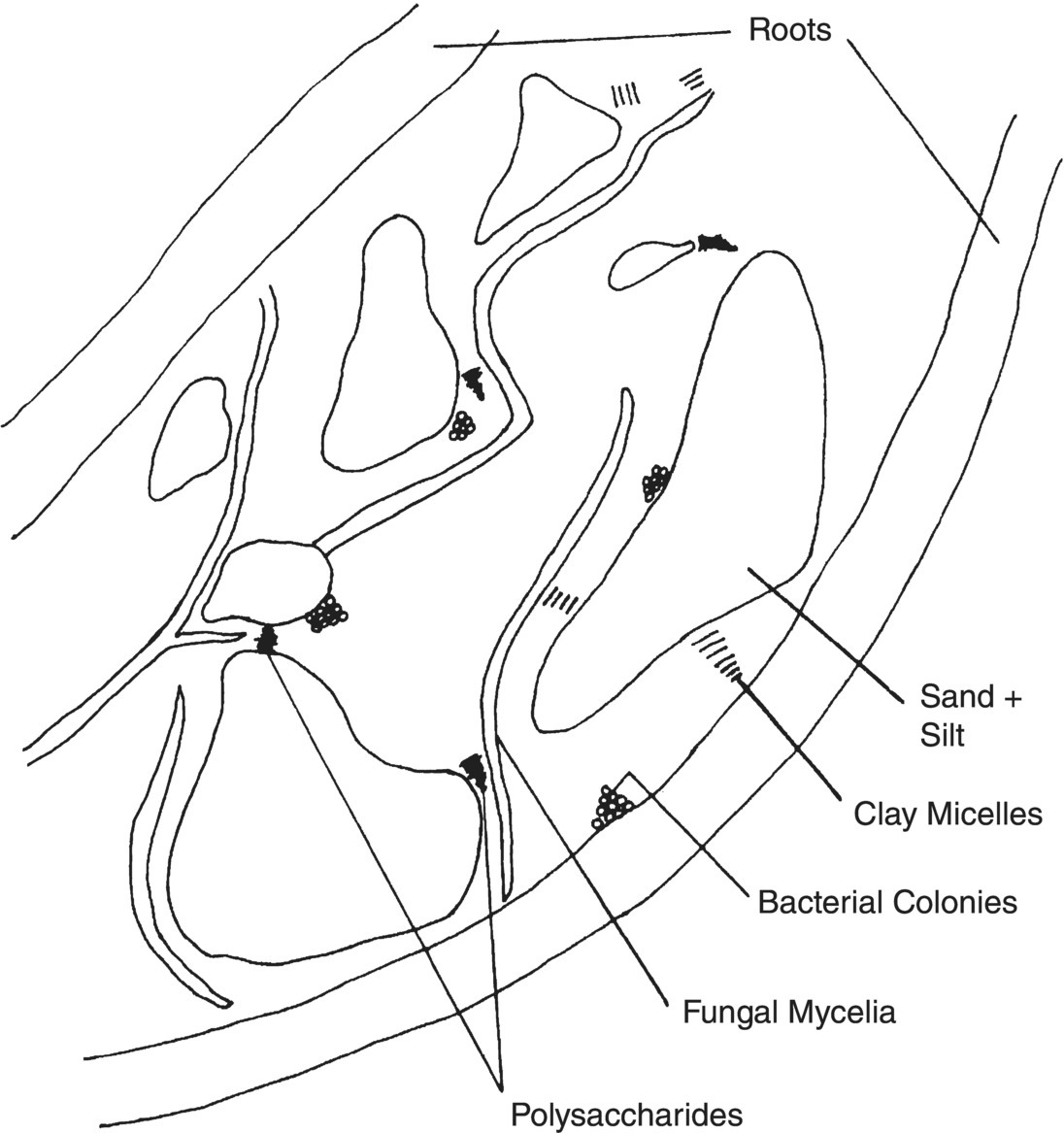
Figure 1.9 Basic elements of soil substituents contributing to aggregate formation (not drawn to scale).
A classic conceptual model presented by Tisdall and Oades (1982) provides insight into the role of the biological community in aggregate formation and a basis for proposing a model of how aggregate formation represents a synergistic interaction between aboveground and belowground life forms. In the model, organic binding agents are divided into three categories: transient (mainly polysaccharides), temporary (roots and fungal hyphae), and persistent (humic substances associated with polyvalent metal cations). Soil aggregates are formed in a hierarchical arrangement by stabilization of smaller aggregates into larger structures by different binding agents (Waters and Oades 1991). Microaggregates are stabilized by persistent binding agents, whereas polysaccharides and biomass (microbial and root) result in the less persistent association of the microaggregates into macroaggregates (Beare et al. 1997; Elliott 1986; Hu et al. 1995). Microaggregates consist primarily of clay and humified organic materials linked by polyvalent metals (Edwards and Bremner 1967). Due to the biodegradation resistance of the persistent binding agents, the microaggregate structure is long term and appears to be more a characteristic of the soil than a product of soil management procedures. In contrast, the assemblage of microaggregates into macroaggregates is less stable and affected more by soil management due to its reliance on soil fungal and root development, including mycorrhizal associations (Hu et al. 1995). This model emphasizes the importance of polysaccharides and humic substances in soil aggregate formation, but it must be appreciated that any organic compound that stimulates or decreases microbial growth in soil indirectly affects aggregate formation by affecting the growth dynamics of the fungal community.
As could reasonably be anticipated by the high variability of soil mineral components and organic matter, the complexity of the micro‐ and macroaggregate structure is more complex than implied by the Tisdall and Oades (1982) model. For example, Watteau et al. (2012) demonstrated that associated with the variable complexity and composition of soil mineral matter comes an increased complexity of classes of microaggregates. The quantity of the organic matter entering the soil affects the dynamics of aggregate formation and stability. For example, more easily decomposable organic residues tend to result in a rapid transient increase in aggregates whereas more slowly mineralized organic matter results in a slower but longer lasting response to the increased energy supply to the soil microbial community, as exemplified by research of Le Guillou et al. (2012). Organic matter quality and its nitrogen content also affect the dynamics of aggregate formation and stability (for example, see Le Guillou et al. 2012 and Gude et al. 2012). Note that the impact of organic matter on this important aspect of soil structure results from turnover of organic matter produced by the native plant community as well as the impacts associated with anthropogenic management of the soil.
Role of carbohydrates in soil aggregate formation: The importance of polysaccharides as binding agents for adhesion of soil mineral components into aggregates has long been appreciated. Early research involved correlation of polysaccharide levels with quantities of water‐stable aggregates, quantifying the effect of periodate oxidation of polysaccharides on aggregation, and analyzing the influence of polysaccharide adsorption on the mechanical strength of clay minerals. For example, Chaney and Swift (1984) found that total organic matter, total carbohydrate, and humic material exhibited high or very high significant correlations with aggregate stability in 120 soils. To rule out other soil physical properties as effectors of aggregate formation, 26 agricultural soils were evaluated. Only organic matter and associated properties correlated significantly with aggregate stability. Other soil parameters examined were sand, silt, clay, iron content, and cation exchange capacity.
As noted above, direct evidence for the role of polysaccharides in soil has been gained by assessing the effect of their selective destruction on soil aggregation. Periodate treatment of soil results in oxidation of polysaccharides contained therein. Not all polysaccharides are destroyed by such treatment. The proportion of polysaccharides oxidized depends on the duration of the treatment and the extent of contact of the polysacharide with the oxidizing agent. The polysaccharide may be occluded by close association within soil particulate structures. Even with these limitations, a major role of polysaccharides in aggregate formation is revealed by this method. Periodate treatment of soil from a long‐term grass field destroyed microaggregates greater than 45 μm in diameter along with the carbohydrates (Cheshire et al. 1983). Residual carbohydrate was enriched in sugars commonly found in plant materials, suggesting preferential destruction of microbial polysaccharides by the periodate. The relationship between soil polysaccharide and aggregate stability was verified in 15 soils and seven soil series (Cheshire et al. 1984).
Chenu and Guerif (1991) assessed the strength of the association between polysaccharides by assessing the mechanical strength of mixtures of kaolinite or montmorillonite plus or minus the polysaccharide scleroglucan. The polysaccharide increased the mechanical strength in mixtures of both clays through the formation of polymer bridges between the clay particles and from the progressive coating of the clay surface by the polysaccharide. The primary effect of the polysaccharide on soil aggregation was therefore the result of increased strength of interparticle bonds.
Humic acids and aggregate stability: Microaggregate formation results from interactions between humic substances and clay particles (for example, see Capriel et al. 1990; Chefetz et al. 2002; Dinel et al. 1991; Schnitzer and Kodema 1992). Aliphatic side chains are particularly instrumental in this process. A high correlation between soil aliphatic fractions (Capriel et al. 1990) and aliphatic‐enriched humic fractions (Dinel et al. 1991) with aggregate stability underscores the importance of these fractions in microaggregate formation. As was noted above, humic acid‐associated aliphatics can form interlayer associations with clays (Theng et al. 1986), which are enriched in soil clay fractions (Arshad and Lowe 1966; Schnitzer et al. 1988). An early study by Schnitzer et al. (1988) demonstrated that although the quantities of aliphatic fraction extracted from soil with supercritical‐hexane varied with management, the chemical properties revealed by 13C NMR and Fourier‐transform infrared differences were independent of management (Schnitzer et al. 1988).
Soil management and aggregate stability: The stability of microaggregates and macroaggregates reflects the biodegradation susceptibility of the primary organic polymers or biological biomass instrumental in their formation. Microaggregates stabilized through association of humic substances with clay particles are more resistant to management procedures and are more reflective of soil physical properties.
Soil management typically results in modification of factors controlling soil microbial activity. For example, cultivation of soil can enhance the degradation of organic matter previously occluded by soil minerals, increases in soil aeration, and modification of soil pH through liming. Portions of the soil organic fractions could be depleted under appropriate conditions in a matter of days or weeks. Therefore, soil management procedures that cause a decrease in soil organic components result in a reduction of quantities of the water‐stable macroaggregates produced by their interaction with soil particulates. Examples of declines in soil structural properties are found in studies of the effect of long‐term cultivation on soil properties. For example, Tiessen and Stewart (1983) evaluated the effect of cultivation for between four and 90 years on soil particle size distribution of three North American grassland soils (Cryoborolls). They noted the continuous depletion of organic matter and the loss of particles >50 μm diameter with time of cultivation, suggesting that the more degradable metabolizable soil organic matter components supporting the structure of soil aggregates were being depleted with time.
Conversely, soil management procedures chosen to increase microbial activity, resulting in an increase in biomass and polysaccharide synthesis, cause improvements in soil aggregate structure, as was reflected in the study of Tisdall and Oades (1982), where the duration and magnitude of the improvement in soil microstructure were shown to be dependent on the nature of the metabolized carbon substrate added to the soil samples. Maximum aggregation occurred in soil amended with the rapidly metabolized substrate, for example glucose, whereas the more lasting but less dramatic improvement in soil structure was detected with more slowly metabolized cellulose. Although cellulose is much more biodegradation resistant than is glucose (months compared to days for depletion from soil, respectively), each of the materials could be considered to be rapidly catabolized when compared with humic substances (degradation times measured in decades or more). Reduction of tillage intervention with crop production is another example of a means of manipulating soil communities and organic matter levels in order to improve aggregate structure. Changing from an intensive cultivation management strategy to no‐till or minimum‐till operation results in increases of soil humus levels and augmented microbial biomass, which coincides with improvements in the soil aggregate structure.
The impact of carbon inputs from crop growth on aggregate formation also can vary with the species grown. For example, Angers (1992), in a comparison of the effect of alfalfa (Medicago sativa L.) or corn (Zea mays L.) growth on the aggregate structure of soils from bare fields, found maximum aggregation under alfalfa with minimal changes in this soil property during the study in the fallow field and that cropped to corn. Most likely, the effect on soil aggregation resulted from a combination of variation in quantities and quality of organic matter available to the soil microbial community plus the resultant alteration of the activity of these populations. This effect of plant growth of soil aggregate structure provides an excellent example of a situation where the aboveground community and soil microbes interact to improve the quality of a degraded soil.
Intensive agriculture resulted in major reductions in the soil organic matter reservoir with a parallel reduction in soil quality and crop yields, thereby having significant impacts on associated communities and societal development (e.g. see French 2010; Kareiva et al. 2007). Appropriate cropping practices can increase soil aggregation and optimize soil microbial activity and crop yields. The increased formation of soil aggregates results in better soil quality, allowing for improved plant growth. With increasing plant biomass production as soil quality improves, more organic matter is provided to the microbes which results in optimization of activity and productivity of both aboveground and belowground communities. Additional stimulation of the soil organic matter pool and associated improvements in soil quality can also be encouraged by recycling of societal organic waste materials. The effect of such amendment on soil aggregate formation can be substantial. For example, Glauser et al. (1988) found an increase of water‐stable aggregates to 85% on soils receiving biosolids amendments, compared to 45% in nonamended soils.
1.3 The Macro‐ecosystem
An appreciation of variation in soil properties on a macro‐scale is equally important to understanding the microsite characteristics described above. Both the vertical and horizontal variation in soil properties limiting microbial activity must be quantified.
Although a considerable interest is developing in estimating microbial function in deep aquifer materials, most soil microbiology research has been aimed at developing an understanding of processes occurring at the point of contact between the aboveground and belowground communities. Thus, soils from the 0, A, and to some extent the upper portions of the B horizon are most frequently studied. Interactions in the B horizon are especially important in sites where the 0 horizon is nonexistent and the A horizon is thin or has been lost through erosion.
The importance of these major surface soil horizons is derived from the fact that the primary energy source for the soil microbial community is photosynthate entering soil primarily as plant biomass or root exudates. Although some water‐soluble plant‐fixed carbon may leach through the soil profile, especially in soils with measurable organic horizons, the primary microbial encounter with organic carbon‐containing materials is in the top few centimeters of the soil profile. Thus, a major concern in assessing the microbial community is the distribution of carbon inputs on the soil surface – that is, plant community growth and dispersion of fixed carbon within the soil profile due to past biomass productivity and its transformation and redistribution within the soil matrix. Secondary considerations relate to variation of physical and chemical growth affecting parameters across the soil surface and throughout the profile. Again, many of these result from activity of the aboveground community. Other factors that must be considered are temperature (which varies diurnally as well as seasonally and is affected by plant density and degree of canopy closure) and moisture (which may also vary seasonally and is affected by topography and plant canopy development). Distribution and amount of microbial activity in soil horizons can also be related to soil parent material and soil developmental history as well as current properties such as pH and salinity. Distribution and range of variation of each of these factors must be considered when extrapolating microbiological data to the ecosystem level.
Variation in soil microbial activity in surface soils is generally controlled by inputs of energy in the form of photosynthetically fixed carbon contained in or exuded from plant biomass. The impact of this energy supply distribution is exemplified by examining the sources of fixed carbon in a temperate forest soil. Within the forest soil, microbial energy is derived from oxidation of fixed carbon contained in litter and from decomposition of root exudates, sloughed cells, and fine roots. The contribution of fine roots to the soil nutrient pool is frequently overlooked, but it can constitute a major source of nutrients and energy for soil microbes (see McClaugherty et al. 1982, 1984; Vogt et al. 1983 for examples). In two temperate forest stands, fine root production was roughly equivalent to leaf biomass production (McClaugherty et al. 1982). If an O horizon has developed, microbial activity in the A horizon is also influenced by fixed carbon carried in leachates and by that portion of the plant biomass mixed into the soil surface by earthworm and other animal activity. Thus, maximal soil microbial activity would be anticipated to occur in the vicinity of the tree or shrub (and grass if it occurs on the forest floor) and under areas of litter accumulation and decomposition.
A similar distribution pattern of plant biomass is observed in a grassland soil. Grass biomass enters soils as leafy material in the thatch and as root exudates and decaying root biomass in a dense root structure. Higher concentrations of water‐stable aggregates are generally found in soils collected from grassland ecosystems compared to soils from other major ecosystem types, if all other limiting factors are equivalent. This results from the high input of biomass, the consistent production of this fixed carbon throughout the growing season, and the proportion of readily decomposable components constituting the plant debris as compared to alternate ecosystem types such as forest or agricultural soils. For example, Jenkinson and Ayanaba (1977) noted that 70% of ryegrass (Lolium multiflora) decomposed with a half‐life of 0.25 years in agricultural soils of Rothamsted, England. The remainder had a half‐life of eight years.
With agricultural soils, the organic matter inputs are commonly similar to those of the grassland in quality, but their distribution and quantities are more limited than is observed with the grassland. Thus, soil structural development is reduced. In intensively cultivated sites, reduction of colloidal organic matter levels in the soil results in a loss of soil microstructure (see Tate 1987 for further analysis of limitations and soil structure in this ecosystem type). Analysis of microbial activity in these soils must be conducted with an understanding of crop planting patterns, cropping history, and tillage practices. In widely spaced grass crops, such as sugarcane (Saccharum spp. L.), biomass inputs to the microbial community between rows are minimal.
Organic matter generally accumulates in swamp ecosystems. Organic matter inputs from the aboveground plant community are limited by the high soil water table, as is the decomposition rate of the decaying plant biomass. The accretion of organic matter indicates that biomass production occurs more rapidly than its decomposition. Other organic inputs to the swamp system originate from erosion of soils. Since the A horizon soils contain most of the colloidal soil organic matter, washing of these soils down slope results in importing of this organic matter into the swamp environment.
1.4 Concluding Comments
The act of allowing a soil sample to flow between one's fingers does not leave the impression that a living, respiring community is functioning therein. It could rather easily be concluded that the utmost importance in the various soil types must be attributed to their physical components – that is, their sand, silt, clay, and, perhaps, rock and pebble contents. Although such a lack of apparent importance of biological entities is at first disappointing, such an observation is critical for the development of our concepts of soil microbiology. It is too easy for soil microbiologists to concentrate on the extremely diverse community of microbes existing in a soil sample and their vast array of genotypic and phenotypic properties, and to forget that these organisms are of no benefit to the total ecosystem if they are incapable of functioning within the soil physical matrix. The physical and chemical properties of soils and their variability (macro‐ and microsite) must always be appreciated if a true understanding of the limitations and capabilities of the soil microbial community is to be fully developed.
The variability of the soil environment and its impact on life processes are both intriguing and frustrating. As microbiologists are keenly aware of the fact that no typical microbe exists for even the simplest physiological or ecological study, they must also appreciate the fact that there is no standard soil. Soil components and their interactions are at least as variable as are the characteristics of their associated microbial populations. Thus, a complete understanding of soil microbial behavior and its contributions to total ecosystem function must include an appreciation of its physical environment – both the mix of individual soil components as well as the physical impediments presented to the soil microbes by the assembly of these discrete particles into a defined soil structure, including horizonal development as well as aggregate formation. Assessment of the soil type and its management history is as important to the conducting of “good science” as is provision of a genus or species name to the microbes studied. That is, a complete soil microbiological study must result not only in an appreciation of the nuances of the biological aspects of the soil ecosystems, but also in a conceptualization of the impact of variations in the physical and chemical environment on the expression of the living capacities.
References
1. Amundson, R. and Jenny, H. (1991). The place of humans in the state factor theory of ecosystems and their soils. Soil Sci. 151: 99–109.
2. Angers, D.A. (1992). Changes in soil aggregation and organic carbon under corn and alfalfa. Soil Sci. Soc. Am. J. 56: 1244–1249.
3. Arshad, M.A. and Lowe, L.E. (1966). Fractionation and characterization of naturally occurring organo‐clay complexes. Soil Sci. Soc. Am. Proc. 30: 731–735.
4. Bartha, R. (1980). Pesticide residues in humus. ASM News 46: 356–360.
5. Beare, M.H., Hus, S., Coleman, D.C., and Hendrix, P.F. (1997). Influences of mycelial fungi on soil aggregation and organic matter storage in conventional and no‐tillage soils. Appl. Soil Ecol. 5: 211–219.
6. Bollag, J.‐M., Liu, S.Y., and Minard, R.D. (1980). Cross‐coupling of phenolic humus constituents and 2,4‐dichlorophenol. Soil Sci. Soc. Am. J. 44: 52–56.
7. Bondietti, E., Martin, J.P., and Haider, K. (1972). Stabilization of amino sugar units in humic‐type polymers. Soil Sci. Soc. Am. Proc. 36: 597–602.
8. Boyd, S.A. and King, R. (1984). Adsorption of labile organic compounds by soil. Soil Sci. 137: 115–119.
9. Brady, N.C. and Weil, R.R. (2017). The Nature and Property of Soils. New Jersey: Prentice Hall.
10. Cao, X., Olk, D.C., Chappell, M. et al. (2011). Solid‐state NMR analysis of soil organic matter fractions from integrated physical‐chemical extraction. Soil Sci. Soc. Am. J. 75: 1374–1384.
11. Capriel, P., Beck, T., Borchert, H., and Harter, P. (1990). Relationship between soil aliphatic fraction extracted with supercritical hexane, soil microbial biomass, and soil aggregate stability. Soil Sci. Soc. Am. J. 54: 415–420.
12. Chaney, K. and Swift, R.S. (1984). The influence of organic matter on aggregate stability in some British soils. J. Soil Sci. 35: 223–230.
13. Chefetz, B., Tarchitzky, J., Deshmukh, A.P. et al. (2002). Structural characterization of soil organic matter and humic acids in particle‐size fractions of an agricultural soil. Soil Sci. Soc. Am. J. 66: 121–141.
14. Chenu, C. and Guerif, J. (1991). Mechanical strength of clay minerals as influenced by an adsorbed polysaccharide. Soil Sci. Soc. Am. J. 55: 1076–1080.
15. Cheshire, M.V., Sparling, G.P., and Mundie, C.M. (1983). Effect of periodate treatment of soil on carbohydrate constituents and soil aggregation. J. Soil Sci. 34: 105–112.
16. Cheshire, M.V., Sparling, G.P., and Mundie, C.M. (1984). Influence of soil type, crop and air drying on residual carbohydrate content and aggregate stability after treatment with periodate and tetraborate. Plant and Soil 76: 339–347.
17. Dec, J. and Bollag, J.‐M. (1997). Determination of covalent and noncovalent binding interactions between xenobiotic chemicals and soil. Soil Sci. 162: 858–874.
18. Dinel, H., Mehuys, G.R., and Levesque, M. (1991). Influence of humic and fibric materials on the aggregation and aggregate stability of a lacustrine silty clay. Soil Sci. 151: 146–158.
19. Duxbury, J.M. and Tate, R.L. III (1981). The effect of soil depth and crop cover on enzymatic activities in Pahokee muck. Soil Sci. Soc. Am. J. 45: 322–328.
20. Edwards, A.P. and Bremner, J.M. (1967). Microaggregates in soil. J. Soil Sci. 18: 64–73.
21. Elliott, E.T. (1986). Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 50: 627–633.
22. Fanning, D.S. and Fanning, M.C.B. (1989). Soil Morphology, Genesis, and Classification. New York: Wiley.
23. Fisk, A.C., Murphy, S.L., and Tate, R.L. III (1999). Microscopic observations of bacterial sorption in soil cores. Biol. Fertil. Soils 28: 111–116.
24. French, C. (2010). People, societies, and landscapes. Science 328: 443–444.
25. Glauser, R., Doner, H.E., and Paul, E.A. (1988). Soil aggregate stability as a function of particle size in sludge‐treated soils. Soil Sci. 146: 37–43.
26. Goring, C.A.I. and Bartholomew, W.V. (1951). Microbial products and soil organic matter: III. Adsorption of carbohydrate phosphates by clays. Soil Sci. Soc. Am. Proc. 15: 189–194.
27. Goring, C.A.I. and Bartholomew, W.V. (1952). Adsorption of mononucleotides, nucleic acids, and nucleoproteins by clays. Soil Sci. 74: 149–164.
28. Greenland, D.J. (1971). Interactions between humic and fulvic acids and clays. Soil Sci. 111: 34–41.
29. Gude, A., Kandler, E., and Gleixner, G. (2012). Input related microbial carbon dynamic of soil organic matter in particle size fractions. Soil Biol. Biochem. 47: 209–219.
30. Haider, K. and Martin, J.P. (1988). Mineralization of 14‐C labelled humic acids and of humic‐acid bound C‐xenobiotics by Phanerochaete chrysosporium. Soil Biol. Biochem. 20: 425–429.
31. Hatcher, P.G., Schnitzer, M., Dennis, L.W., and Marciel, G.E. (1981). Aromaticity of humic substances in soils. Soil Sci. Soc. Am. J. 45: 1089–1094.
32. Hu, S., Coleman, D.C., Beare, M.H., and Hendrix, P.F. (1995). Soil carbohydrates in aggrading and degrading agroecosystems‐influences of fungi and aggregates. Agric. Ecosyst. Environ. 54: 77–88.
33. Hwang, S. and Tate, R.L. III (1997). Interactions of clay minerals with Arthrobacter crystallopoietes: starvation, survival and 2‐hydroxypyridine catabolism. Biol. Fertil. Soils 24: 335–340.
34. Jenkinson, D.S. and Ayanaba, A. (1977). Decomposition of carbon‐14 labeled plant material under tropical conditions. Soil Sci. Soc. Am. J. 41: 912–915.
35. Jenny, H. (1941). Factors of Soil Formation; A System of Quantitative Pedology. New York: McGraw‐Hill Book Co.
36. Kareiva, P., Watts, S., McDonald, R., and Boucher, T. (2007). Domesticated nature: shaping landscapes and ecosystems for human welfare. Science 316: 1866–1869.
37. Kodema, H. and Schnitzer, M. (1974). Adsorption of fulvic acid by non‐expanding clay minerals. Trans. Int. Congr. Soil Sci. 10 (2): 51–56.
38. Kunc, F. and Stotzky, A.G. (1974). Effect of clay minerals on heterotrophic microbial activity in soil. Soil Sci. 118: 186–195.
39. Le Guillou, C., Angers, D.A., Maron, P.A. et al. (2012). Linking microbial community to soil water‐stable aggregation during crop residue decomposition. Soil Biol. Biochem. 50: 126–133.
40. Li, L., Huang, W., Peng, P. et al. (2003). Chemical and molecular heterogeneity of humic acids repetitively extracted from a peat. Soil Sci. Soc. Am. J. 67: 740–746.
41. Lichtenstein, E.P. (1980). “Bound” residues in soils and transfer of soil residues in crops. Residue Rev. 76: 147–153.
42. Liu, S.‐Y., Greyer, A.J., Minard, R.D., and Bollag, J.‐M. (1985). Enzyme‐catalyzed complex‐formation of amino acid esters and phenolic humus constituents. Soil Sci. Soc. Am. J. 49: 337–342.
43. Lynch, D.L. and Cotnoir, L.J. Jr. (1956). The influence of clay minerals on the breakdown of certain organic substrates. Soil Sci. Soc. Am. J. 20: 367–370.
44. Malcolm, R.L. (1990). The uniqueness of humic substances in each of soil, stream, and marine environments. Anal. Chem. Acta 232: 19–30.
45. McClaugherty, C.A., Aber, J.D., and Melillo, J.M. (1982). The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. Ecology 63: 1481–1490.
46. McClaugherty, C.A., Aber, J.D., and Melillo, J.M. (1984). Decomposition dynamics of fine roots in forested ecosystem. Oikos 42: 378–386.
47. Nayak, D.C., Varadachari, C., and Ghosh, K. (1990). Influence of organic acidic functional groups of humic substances in complexation with clay minerals. Soil Sci. 149: 268–272.
48. Novak, J.M. and Smeck, N.E. (1991). Comparisons of humic substances extracted from contiguous Alfisols and Mollisols in Southwestern Ohio. Soil Sci. Soc. Am. J. 55: 96–102.
49. Olnes, A. and Clapp, C.E. (1972). Microbial degradation of a montmorillonite‐dextran complex. Soil Sci. Soc. Am. J. 36: 179–181.
50. Piccolo, A., Conte, P., Trivellone, E. et al. (2002). Reduced heterogeneity of a lignite humic acid by preparative HPSEC following interaction with an organic acid. Characterization of size‐separates by Pyr‐GC‐MS and 1H‐NMR spectroscopy. Environ. Sci. Technol. 36: 76–84.
51. Preston, C.M. (1996). Applications of NMR to soil organic matter analysis: history and prospects. Soil Sci. 161: 144–166.
52. Saxena, A. and Bartha, R. (1983a). Microbial mineralization of humic acid‐3,4‐dichloroaniline complexes. Soil Biol. Biochem. 15: 59–62.
53. Saxena, A. and Bartha, R. (1983b). Binding of 3,4‐dichloroaniline by humic acid and soil: mechanism and exchangeability. Soil Sci. 136: 111–116.
54. Schnitzer, M. and Kodema, H. (1992). Interactions between organic and inorganic components in particle‐size fractions separated from four soils. Soil Sci. Soc. Am. J. 56: 1099–1105.
55. Schnitzer, M. and Preston, C.M. (1986). Analysis of humic acids by solution and solid‐state carbon‐13 nuclear magnetic resonance. Soil Sci. Soc. Am. J. 50: 326331.
56. Schnitzer, M., Ripmeester, J.A., and Kodama, H. (1988). Characterization of the organic matter associated with a soil clay. Soil Sci. 145: 448–454.
57. Schulten, H.‐R. and Schnitzer, M. (1997). Chemical model structures for soil organic matter and soils. Soil Sci. 162: 115–130.
58. Schulten, H.‐R. and Schnitzer, M. (1990). Aliphatics in soil organic matter in fine‐clay fractions. Soil Sci. Soc. Am. J. 54: 98–105.
59. Simpson, M.J. (2006). Nuclear magnetic resonance based investigations of contaminant interactions with soil organic matter. Soil Sci. Soc. Am. J. 70: 995–1004.
60. Sjoblad, R.D. and Bollag, J.‐M. (1981). Oxidative coupling of aromatic compounds by enzymes from soil microorganisms. In: Soil Biochemistry, vol. 5 (eds. E.A. Paul and J.N. Ladd), 113–152. New York: Marcel Dekker.
61. Sorensen, L.H. (1972). Stabilization of newly formed amino acid metabolites in soil by clay minerals. Soil Sci. 114: 5–11.
62. Sorensen, L.H. (1975). The influence of clay on the rate of decay of amino acid metabolites synthesized in soils during decomposition of cellulose. Soil Biol. Biochem. 7: 171–177.
63. Stearman, G.K., Lewis, R.J., Tortorelli, L.J., and Tyler, D.D. (1989). Characterization of humic acid from no‐tilled and tilled soils using carbon‐13 nuclear magnetic resonance. Soil Sci. Soc. Am. J. 53: 744–749.
64. Stevenson, F.J. (1994). Humus Chemistry. Genesis, Composition. Reactions. New York: Wiley.
65. Still, C.C., Hsu, T.‐S., and Bartha, R. (1980). Soil‐bound 3,4‐dichloroaniline: source of contamination in rice grain. Bull. Environ. Contam. Toxicol. 24: 550–554.
66. Tate, R.L. III (1987). Soil Organic Matter. Biological and Ecological Effects. New York: Wiley.
67. Theng, B.K.G., Churchman, G.J., and Newman, R.H. (1986). The occurrence of interlayer clay‐organic complexes in two New Zealand soils. Soil Sci. 142: 262266.
68. Tiessen, H. and Stewart, J.W.B. (1983). Particle‐size fractions and their use in studies of soil organic matter: II. Cultivation effects on organic matter composition in size fractions. Soil Sci. Soc. Am. J. 47: 509–514.
69. Tisdall, J.M. and Oades, J.M. (1982). Organic matter and water‐stable aggregates in soils. J. Soil Sci. 33: 141–163.
70. Vogt, K.A., Grier, C.C., Meier, C.E., and Keyes, M.R. (1983). Organic matter and nutrient dynamics in forest floors of young and mature Abies amabilis stands in Western Washington, as affected by fine‐root inputs. Ecol. Monogr. 52: 139–157.
71. Von Wandruszka, R. (1998). The micellar model of humic acid: evidence from pyrene fluorescence measurements. Soil Sci. 163: 921–930.
72. Waters, A.G. and Oades, J.M. (1991). Organic matter in water‐stable aggregates. In: Advances in Soil Organic Matter Research: The Impact on Agriculture and the Environment (ed. W.S. Wilson), 163–174. Cambridge: Royal Society of Chemistry.
73. Watteau, F., Villemin, G., Bartoli, F. et al. (2012). 0–20 μm aggregate topology based on the nature of aggregative organic materials in a cultivated silty topsoil. Soil Biol. Biochem. 46: 103–114.