Chapter Eleven
INTRODUCTION
Like many other biological systems, coral reef communities are subjected to a variety of environmental disturbances of varying intensity, frequency and duration (Pearson 1981; Huston 1985; Karlson and Hurd 1993). The nature and magnitude of impacts associated with the various natural disturbances, therefore, also vary with the nature and intensity of the disturbance. Tropical coral seas have often been regarded in the past as relatively stable environments (Johannes 1975; Endean 1976), without pronounced seasonal environmental fluctuations (e.g., temperature). In comparison, wide environmental fluctuations (e.g., temperature) are a rule in most shallow-water environments of the higher latitudes.
The environmental stability assumption is generally true, especially within tropical archipelagic seas, but many areas of the tropical oceans are also subjected to "short-term" unpredictable events like cyclones, volcanic eruptions, earthquakes and tsunamis. As a result of differing viewpoints, with regards to spatial and temporal scales, the idea that tropical regions, specifically coral reef environments, are stable systems has been severely criticized (Grigg and Dollar 1991). When viewed from a time scale of millions of years (i.e., the geological or evolutionary time frame), these seemingly unpredictable short-term disturbances may be regarded as components of a relatively stable system. However, it has been shown that short-term unpredictable disturbances, or the absence of disturbances, when viewed from a time scale of centuries or decades (i.e., the ecological time frame), are the principal driving forces in the structuring of recent coral reef communities.
ATMOSPHERIC DISTURBANCES
The fact that the archipelago lies outside the Indo-Pacific cyclone belt (fig. 11.1) may have had a pronounced influence on the evolutionary history of Recent reef-building corals, and therefore, on the function and structure of coral reef ecosystems in this area. Long-term environmental stability, coupled with high habitat diversity (on both local and regional scales) (Best and Boekschoten 1988), and an abundant supply of energy and nutrients, are the key features favoring highly diverse and "biologically controlled" (Sanders 1968; Porter 1972a,b, 1974) coral reef communities.
Structural complexity and zonation patterns of coral communities along the southern regions of the archipelago, exposed to the swell of the Indian Ocean, are distinctly different from those in the more protected archipelagic seas. Coral communities along the southern coastline of many East Nusa Tenggara islands are also subjected to heavy seas from frequent cyclones that form in the Indian Ocean and Timor Gap, just off the western coast of Australia. To the north, the reefs along the north coast of Irian Jaya, Halmahera, and the Sangihe and Talaud Archipelagoes are exposed to the swell generated by distant, and not-so-distant, tropical cyclones passing through the Philippines.
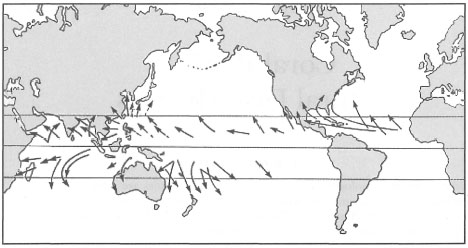
Figure 11.1. General tracts of tropical cyclones in the Indo-Pacific region, illustrating that the Indonesian Archipelago lies outside the cyclone belt.
From Open University 1991b. Circulation p. 27; fig. 2.15.
Unfortunately, comparative studies between these different environments have not been conducted. However, recent work by Dr. C. Wallace, from die Museum of Tropical Queensland, who has been studying the distribution of Acropom in Indonesia, has resulted in some exciting new discoveries concerning their distribution, environmental conditions, and not surprisingly, numerous new species.
Surface winds throughout the archipelago can be classified as generally light. Normal wind speeds along the coastlines are usually between 7-11 km.hr most of the year, and considerably lower inland (RePPProT 1990). On most large islands, land and sea breezes are a dominant and predictable phenomena that have served the coastal fishermen for centuries.
Monsoonal Storms
The impacts of winds generated by localized storms (e.g., squalls) are qualitatively and quantitatively different from those of tropical cyclones. Strong winds may sometimes reach gale-force strength (Beaufort No. 8-9; 45-65 km.hr-1), but these are very rare and occur primarily in the upland areas of large islands (e.g., Sumatra, Kalimantan, Java, etc.). Strong wind gusts are common, especially between March April and September-October, which roughly coincides with the equinoxes (RePP-ProT 1990). There have been reports of occasional tropical revolving storms, and evidence of tornado damage has been reported from Irian Jaya. Occasionally, severe tropical storms may be associated with intense rainfall, which at times may reach >700 mm.day , posing considerable safety hazards to large structures such as bridges (RePPProT 1990).
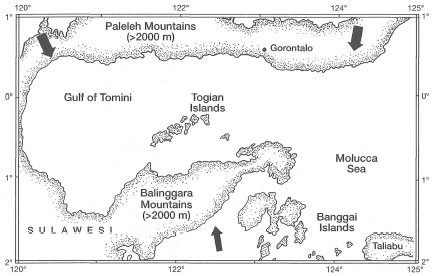
Figure 11.2. A map of the Gulf of Tomini illustrating the sheltered nature of the Togian Islands with extensive reef development. The arrows from the north and south indicate predominant wind directions during the Northwest and Southeast Monsoons, respectively.
While some storms can generate almost cyclone-force winds, the duration and fetch are insufficient for the establishment of heavy sea conditions, which rarely exceed 3 m amplitude. Most severe storms seem to occur in the shallow Java Sea, which has a relatively long fetch-length along an east-to-west axis. The influence of the steady Southeast Monsoon winds on the geomorphology of the Kepulauan Seribu Reefs is well-known (Molengraaff 1928; Kuenen 1933; Umbgrove 1948).
Because of the seasonality and relative predictability of the monsoons, coral communities within the archipelagic seas exposed to monsoonal winds have developed characteristic zonation patterns that differ greatly from those in sheltered environments. However, the geological complexity, on regional and local scales, offers a variety of exposed and sheltered shallow-water habitats that modify the influence of atmospheric disturbances. The highly diverse coral reefs (including fringing reefs, barrier reefs and atolls) of the Togian Islands, in the Gulf of Tomini, are a good example of a sheltered system. The Paleleh Mountains to the north (i.e., the North Arm of Sulawesi) and the Balinggara and Batui Mountains to the south (i.e., the East Arm of Sulawesi) provide an effective protection against the winds of the Northwest and Southeast Monsoons respectively (fig. 11.2).
Box. 11.1. Local weather phenomena over the Indonesian Archipelago.
P. A. Winarso, Meteorological and Geophysical Agency of Indonesia, Jakarta, Indonesia.
The Indonesian Archipelago occupies a large equatorial region between 06° N and 12° S and 90° E to 150° E. Land represents only about 33% of the total area of the archipelago, thus Indonesia is a maritime nation. The Indonesian Archipelago is under the influence of the Maritime Continental Air Mass, resulting in a humid and warm climate. Because of the characteristics of this air mass and convective activity, clouds are easily generated over this region, causing frequent local storm activity. The effects of local storm events are mainly due to increased turbulence associated with strong gusts of wind. Each year, localized storms, associated with the formation of cumulonimbus clouds, are responsible for environmental damage to shallow-water coastal and marine communities as well as to land.
On average, local storms on land and sea are responsible for more damage to the environment and property than any other natural phenomenon (i.e., earthquakes or volcanic eruptions). Local storms may cause storm surges and high seas that can severely impact upon coastal structures, causing severe damage. Heavy seas associated with strong gusts of wind may also impact on benthic communities in coastal areas as well as deeper water offshore.
Some of the most damaging storms are associated with the formation of highly convective cells known as the cumulonimbus clouds. The cumulonimbus clouds can form rapidly over open seas, with the base of the cloud usually a few hundred metres above sea level, while the top of the clouds can reach altitudes of 15,000 m above sea level. Wind velocities associated with these events, also known as squalls, can reach gale force for brief periods, causing considerable damage, mainly above water. Because of their short duration they do not seem to have a significant impact on coral communities in general, however, exceptions may occur. These storms are most frequent between November and April, and are closely linked to the position of the Inter-Tropical Convergence Zone (ITCZ). In the ITCZ, air masses from the Northern and Southern Hemispheres converge as a result of high surface temperatures. The ITCZ is characterized by frequent storms with heavy rainfall and strong winds.
In addition to the winds associated with the convective cloud activity, strong winds also predominate during the tropical cyclone season of both hemispheres. During these periods local storm surges occur in Indonesian regions that are close to the disturbances. For example, during the tropical cyclone season in the Northern Hemisphere (i.e., May - October), several coastal regions in North Sumatra, East Kalimantan and North Sulawesi may experience storm surges associated with passing cyclones. During the Southern Hemisphere cyclone season, between November - April, coastal areas of Java, Bali, Nusa Tenggara, South Sulawesi and East Timor may experience strong winds associated from the peripheral influence of tropical cyclones in the Southern Hemisphere.
The western regions of the archipelago are also influenced by periodic cold surges originating to the north of the South China Sea. The cold surges occur from November to March, with surface winds that can reach up to 60km.hr-1, creating heavy sea conditions in the Natuna and Java Seas. Heavy seas associated with the cold surges cause considerable damage along the southeast coast of Sumatra, the Riau Islands and the north coast of Java. The storm conditions can be intensified by the presence of the ITCZ, resulting in heavy rainfall and strong, gale-force winds over the shallow Java Sea.
The reefs around the western Togian Islands are situated in a zone, famous for the unruffled calm and undisturbed quiet of the sea… Neither on the barrier reefs, nor on the atolls, nor on the fringing reefs of the Togian Islands does one single shingle rampart occur. In vain I searched for Montipora foliosa,…— UMBGROVE 1947
The protection from the Northwesterly and Southeasterly Monsoonal winds offered by the two mountain ranges has a pronounced effect on the overall geomorphology of Togian reefs. This is especially so in the northwest region of the island chain, since the islands themselves (elevation >500 m) provide additional protection from the influence of strong Southeast Monsoon winds. The most noticeable effect is on the reef zonation as well as on coral community composition. For example, Montipora foliosa was recognized by Umbgrove (1947) as a conspicuous component on the exposed reef slope of Nyamuk Besar (Leiden), a coral cay in Jakarta Bay. Coral communities on Nyamuk Besar are under strong monsoonal influence. M. foliosa was especially abundant along the northwest and southeast slopes of the shingle rampart, demonstrating clear preference for high-energy environments. The northwest and southeast slopes of the reef face the prevailing Northwest and Southeast Monsoon winds, respectively (fig. 11.3).
M. foliosa is also an abundant species (encrusting morphology) throughout the Kepulauan Seribu (i.e., Thousand Islands), especially on the shallow exposed reef slopes. Suharsono (1992) found that M. foliosa was one of the 30 coral species present in all of his sampling stations at Pulau Genteng, Thousand Islands, out of a total of 180 scleractinians. Yet this species is apparently absent from the protected shallow reef slopes of the Togian reefs, even though it is found in deeper water (Umbgrove 1947).
In contrast, the delicate hispidose Acropora turaki (Wallace 1994) was a dominant species along the southern coast of Pulau Talatakoh, with extensive lagoonal environments protected by a barrier reef. In fact, the calm waters of the Togian Islands seem to be an optimal environment for delicate Acropora species (e.g., Acropora jacquelineae [Wallace 1994]; A. lokani [Wallace 1994]; A. caroliniana Nemenzo) which dominated the shallow reefs as well as offshore atoll environments (C. Wallace and J. Wolstenholme, pers. comm.).
In contrast to both Kepulauan Seribu and, the Togian Islands, the oceanic reefs in the Banda Sea (e.g., Nil Desperandun) under oceanic conditions have a much different community. The shallow reef flat and reef slopes are dominated by extensive and dense stands of Acropora palifera, A. cuneata, A. brueggemanni, A. monticulosa and especially A. robusta. The presence of this particular assemblage of acroporids tells us that Nil Desperandun is a high-energy environment subjected to strong wave action and currents. Indeed, during our survey of the reef, current velocity was in excess of 2 m.sec-1, unusually strong for an oceanic platform reef with a tidal amplitude of about 1.5 m.
This superficial look at three different environments suggests that atmospheric disturbances other than tropical cyclones play an insignificant role in the structuring of the shallow-water coral communities in the Togian Islands. Furthermore, the lack of shingle ramparts or coral cays in the region suggests that wind and waves may have negligible roles in the geomorphology of the reef structures. Clearly, this unique region, with an amazing diversity of reef habitats and geomorphologies, deserves a lot more attention from the scientific community than it has received thus far. The coral communities in Kepulauan Seribu, and in the past, in Jakarta Bay as well, show a classical zonation structure that developed in response to predictable monsoonal weather patterns. The coral community on Nil Desperandun indicates that large-scale disturbances may occur.
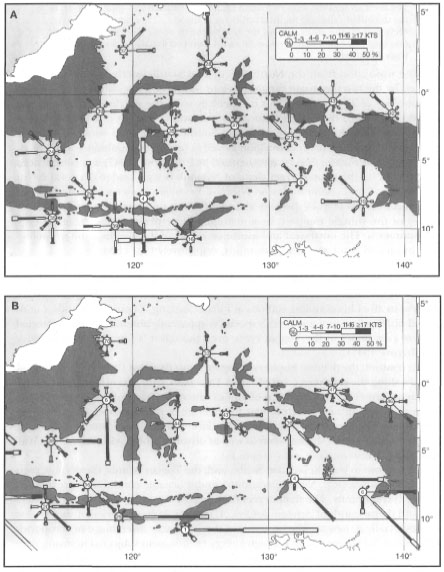
Figure 11.3. Wind roses for the central and eastern archipelago during January (A) and July (B).
Peta Cuaca Perairan Indonesia 1992.

Figure 11.4. Frequency of tropical cyclones in the Indo-Pacific region.
Modified from Scoff in 1993.
Tropical Cyclones
On the numerous reefs of the Great Barrier Reef, one can see large coral boulders that were thrown upon the reefs by rough seas during numerous tropical cyclones. Their noticeable absence from the Indonesian reefs that are nonetheless heavily influenced by strong monsoonal winds has been noted previously (Kuenen 1933; Umbgrove 1947). Based on their absence, one can arrive at the conclusion that tropical cyclones do not occur in the archipelagic seas. This generalization does actually apply to most of the vast archipelag;, however, as was mentioned earlier, the southern islands of East Nusa Tenggara, and the northern islands from Biak to Sangihe, have been known to be affected by cyclonic conditions resulting from a bypassing cyclone. Figure 11.4 gives us some idea on the frequency of cyclones that pass in close proximity to the northern and southern regions of the archipelago.
Figures 11.1 and 11.4 clearly demonstrate that the Indonesian Archipelago lies between two main tropical cyclone regions. However, because of Indonesia's geographic position and prevailing atmospheric conditions, most tropical cyclones move away from the archipelago. Periodically, however, cyclones do wander in, and with rather devastating impacts to the coral communities. Figure 11.5 shows the general tracks of cyclones that have, in the past, strayed into the eastern regions of the archipelago.
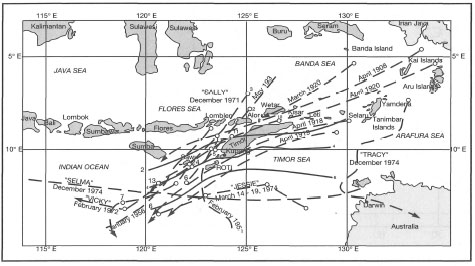
Figure 11.5. Tracks of cyclones in East Nusa Tenggara and Moluccas from 1908 to 1974.
From RePPProT 1990.

Figure 11.6. Tropical cyclone tracts on the Great Barrier Reef and Western Australia from 1908 to 1981.
Courtesy of T. Done and the Australian Institute of Marine Science, Townsville.
There seems to be a well-defined "cyclone corridor" around Timor. Some reports suggest that cyclonic conditions have been recorded as far west as Lombok (RePPProT 1990). The impact of these storms has not been documented previously, and therefore, very little is known about their effect on the coral reef community. If the cyclones are of sufficient strength, we would expect some coral boulders to be thrown upon the reef flats of, at least, the most exposed reefs. However, as was also noted by Umbgrove (1947), corroborative evidence from the islands around Timor is not available. In contrast to their relatively infrequent visits to the eastern part of the archipelago, cyclones are a yearly occurrence on the Great Barrier Reef (fig. 11.6).
CYCLONE "LENA" (JANUARY 23, 1993)
Maumere Bay is renowned not only for the world-class diving, but also for an amazing diversity of coral reef fish. Because of the high diversity of physiographic features found in Maumere Bay, reef diversity is correspondingly high (i.e., fringing reefs, steep drop-off reefs, a barrier reef, and an offshore atoll). As a result of high habitat diversity, the reefs of Maumere Bay support diverse assemblages of corals. Unfortunately, little work has been done on coral community structure in this area, and therefore little information is available on their diversity. However, the numerous reef habitats support an amazing diversity of reef fish fauna, with over 1700 species identified. (G. Allen, pers. comm.).
Figure 11.5 illustrates that tropical cyclones are a relatively rare occurrence in the eastern regions of the archipelagic seas. The last cyclone to pass through the region was "Sally", in December 1971. According to the weather reports, the eye of Cyclone "Lena" did not actually enter the Indonesian seas, however, Flores and the adjacent islands to the east were affected by the winds at the outer periphery of the disturbance. Winds blowing predominantly from the north-northwest (330-350°) reached gusts of up to 45 knots (Beaufort 9 or 80 km.hr -1) (Wai-Oti Meteorological Station, Maumere). Considerable structural damage (e.g., shingles and slate removed from roofs) was reported for properties along the coastline. Unfortunately for many of the shallow-water coral communities, the storm occurred within two months of a major earthquake, which caused severe structural damage to many coral reefs in the area. The extent of cyclone damage in the affected regions is not known, but an example from Maumere Bay may serve to illustrate the destructive power of these disturbances, especially in regions where they rarely occur. Figure 11.9 summarizes our qualitative observations on the impact of Cyclone "Lena" on coral communities at various locations in Maumere Bay.
Consistent with the direction of prevailing winds, the least damage to the coral communities occurred on reefs located in relatively sheltered areas along the southeast coast of Pulau Besar and Pulau Dambilah. The most severe damage occurred along the south shore of Maumere Bay, which was fully exposed to the storm. However, the area was also struck by the December 1992 tsunami. Which disturbance caused more damage to coral communities is difficult to ascertain. Observations of well-sorted beach deposits indicate that the storm disturbance may have had a greater impact than the 2-4 metre-high tsunami. Rough weather conditions apparently persisted for 2-3 days, during which time a significant shingle rampart developed (1-2 m above sea level) along many sections of the northern coastline of Maumere Bay. Well-sorted shingle ramparts are usually constructional features associated with storm events (Umbgrove 1947), and not with tsunamis.
Box 11.2. Tropical Cyclone "Lena", January 1993.
P. A. Winarso; Meteorological and Geophysical Agency of Indonesia, Jakarta, Indonesia.
During January - February 1993, a large-scale atmospheric disturbance occurred over most of the Indonesian Archipelago, extending from the South China Sea through the Java and Flores Seas, to the Arafura Sea. This large-scale atmospheric disturbance was characterized by exceptionally rough seas throughout this region, resulting in the sinking of a number of ships, damage to several oil platforms in the Java Sea, as well as damage to shallow-water coral communities from Java to Flores. Detailed analysis of daily weather reports indicate that two low pressure systems were established in the region, one just northwest of Australia and one over the Gulf of Carpentaria (figs. 11.7 and 11.8).
The northwestern depression strengthened into Cyclone "Lena", which moved with a trochoidal-motion track, initially in a westerly direction. "Lena" was, however, deflected from the normal southwesterly direction eastwards. During this time, the area was also experiencing cold surges that are active in January. Associated with the cold surges were strong southerly winds of up to 30 knots (56 km.hr -1) over the South China Sea, which generated fast southerly surface currents of more than 3 m.s-1 (10.8 km.hr -1), and waves of up to 3 m in amplitude. Similar conditions were observed in the west Java Sea (Bangka Island), where d (3 m.s-1), 10-m-deep surface currents and waves of more than 4 m height were recorded. Associated with this atmospheric disturbance were heavy rains of up to 100 mm along most of the north coast of Java and the islands of Nusa Tenggara (i.e., from Bali to Timor).
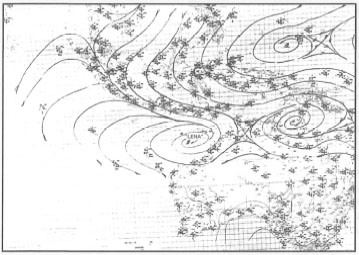
Figure 11.7. Surface weather analyses on 25 January 1993 at 00.00 UTC.
Review of historical weather data indicates that these conditions are infrequent, and thus this event may be classified as a rare occurrence, which poses a question as to what may have caused it. It is suggested that the disturbance was a result of the synergistic effect of the abnormal track of Cyclone "Lena" that coincided with the activity of the cold surge from the South China Sea waters. Normally, tropical cyclones in this region move in a southwesterly direction, and the winds that are generated along the southern part of the Indonesian Archipelago are less than 30 knots. Furthermore, this disturbance followed the "El Niño - Southern Oscillation" (ENSO) event of 1991-1992, which incidentally coincided with the peak sunspot activity (11 -year cycle), characterized by increasing activity of solar flares. It is postulated that if post-ENSO periods coincide with increased surface warming due to increased intensity of solar flares, these types of atmospheric events may occur at infrequent intervals.
It is expected that meteorological and oceanographic conditions associated with tropical cyclones will continue to affect some parts of the Indonesian Archipelago from time to time. Historical review of large-scale weather patterns and oceanographic conditions may allow us to make future predictions on the possibility of occurrence of these rare events in the archipelago.

Figure 11.8. Satellite cloud photograph of Cyclone "Lena" on 25 January 1993 at 00.00 UTC.
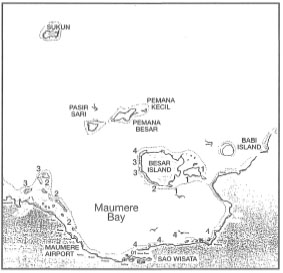
Figure 11.9. Map of Maumere Bay and the location of survey sites in November 1993. The survey applies only to coral communities at depths between 2-15 m. Numbers indicate severity of cyclone damage: 1) Low numerous branching and laminar Acropora spp., abundance of recruits; 2) Medium - visible damage to branching species, massive species present, numerous recruits; 3) High - branching species totally destroyed, massive species damaged but still in place, substrate of unconsolidated rubble and loose sediment is inhibiting recruitment; 4) Severe - most corals destroyed (95%-100%), very few recruits. Recruitment estimate is based on the abundance of juvenile colonies (<10 cm diameter).
Heavy rainfall during and subsequent to the disturbance had an additional impact on most of the coral communities along the south shore of Maumere Bay. The most affected were coral communities on fringing reefs, as well as on the steep drop-offs, located in close proximity to small rivers and seasonal streams. Figure 11.10 demonstrates that rainfall during February 1993 was significantly higher than in January or March 1993.
The fringing reef in front of Flores Sao Wisata Resort, Maumere Bay, is an excellent example of a land-runoff impacted reef. Located to the west of the fringing reef (about 100-200 m) is a small stream with a relatively heavy sediment load (i.e., water full of silt). The runoff from the stream is usually directed away from the reef by a small sand bar and carried westward by the longshore current away from the fringing reef. It seems that the original sand bar was destroyed during the storm by heavy seas impacting the coastline from the north-northwest. As a result of prevailing sea conditions, a new sand bar was created on the western side of the stream, thus directing the stream-runoff towards the fringing reef. Subsequent runoff associated with heavy rains in February smothered most of the reef by fluvial sediments. During our survey in November 1993, the reef sediments were 75%-90% fluvial in origin. Numerous erect, but dead, colonies were covered by a fine layer of volcanic silt and detritus. No recruitment was observed, mainly as a result of fine silt covering most of the available substrate. Reef recovery is not expected until the direction of the stream's discharge is changed to its original position, which should significantly reduce the influx of fluvial sediments, especially during heavy rains.
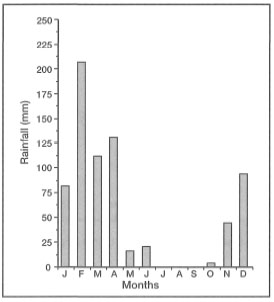
Figure 11.10. Monthly average rainfall (mm) during 1993 in Maumere, Flores, measured at Wai-Oti Meteorological Station. The high rainfall recorded in February is associated with Cyclone "Lena".
EL NINO-SOUTHERN OSCILLATION (ENSO)
It is well-known that the El Niño-Southern Oscillation (ENSO) events are linked closely to the position of an atmospheric low pressure system commonly referred to as the Indonesian Low (fig. 11.11).
The Indonesian Low is located over the archipelago during non-ENSO years, and is characterized by rising, humid, tropical air masses associated with heavy rainfall. The atmospheric low is also closely associated with a western Pacific warm water pool whose mobility is directly linked with ENSO events. During non-ENSO years, steady southeast Trade Winds pile up the warm Pacific Ocean surface waters against the northeast margin of the archipelago, which creates a pressure gradient that allows the warm surface waters from the Pacific Ocean to flow into the archipelagic seas, and into the Indian Ocean (Wyrtki 1961, 1987). During ENSO years, the Indonesian Low becomes a weak low (i.e., atmospheric pressure increases) and is displaced eastwards with a corresponding drop in atmospheric pressure in the South Pacific High. These atmospheric changes cause the western Pacific Ocean warm water pool to flow in an easterly direction. It is, therefore, reasonable to assume that severe ENSO events may also have a measurable effect on sea surface temperature distribution in the archipelagic seas by causing changes in circulation patterns as well as in the position of the thermocline. Whether changes in ocean dynamics associated with ENSO events have an impact, negative or positive, on marine ecosystems in the archipelago is, however, not known. It is interesting to point out, however, that during ENSO events, the thermoclines in both the Timor and Flores Seas rise and sea levels drop (Bray et al. 1994). How these changes impact upon benthic marine communities, especially the intertidal coral reef communities, needs to be addressed in future studies.
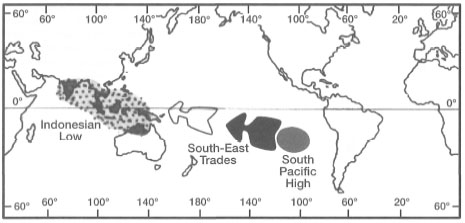
Figure 11.11. The position of the Indonesian Low and the South Pacific High during non-ENSO years.
Modified from Open University 1991b.
El Niño and Coral Reef Bleaching
Whether or not the coral bleaching complexes of the 1980s have been caused by global warming is a critically important yet currently unresolved question. However, the evidence for numerous recurrent, large-scale coral bleaching and mortality events in recent years is unequivocal. Like the destruction of tropical rain forests, the world's coral reef ecosystems are facing major disruptions.—GLYNN 1991
What is coral reef bleaching? What causes coral reef bleaching? Is global warming a threat to coral reefs? These are questions of global significance, that have engaged some of the best minds in coral reef science. Future environmental impacts on coral reefs, associated with climate change, are a major concern of various international institutions and conservation groups as well as national governments (D'Elia et al. 1991). Considering the fact that a great majority of the natural resources associated with coral reef ecosystems fall within the jurisdictions of several developing nations (where they are a key economic asset), the potential repercussions of anthropogenically-induced global climate change are alarming. As has been stated earlier, from the coral management standpoint, it is not whether the coral reefs will survive, but rather whether they will be able to support the resource base necessary for the continuous welfare of the coastal communities that depend on them. Recent mass bleaching events, of still-uncertain origins, have demonstrated that coral reefs can shift from an animal-symbiont-dominated system, which supports a wealth of resources, to a macroalgal-dominated system with significantly lower resource diversity.
Before proceeding any further, however, a clear definition of what is 'coral bleaching' is in order, since a number of purely behavioral responses to natural environmental stressors can be easily diagnosed as bleaching (Brown and Le Tissier 1992). Following the recommendation of Williams and Bunkley-Williams (1990), bleaching is defined as: "Loss of photosymbiotic microorganisms (dinoflagellates, red and green algae, or cyanobacteria), or the pigments of these photosymbionts, or some of both, from tissues of host cnidarians, sponges, mollusks or other photosymbiotic host animals." The definition should also include endosymbiotic diatoms that live in symbiosis with large benthic reef-associated foraminiferans, such as Amphistegina gibbosa (Hallock et al. 1992). The term coral bleaching is now widely accepted as suggested by Williams and Bunkley-Williams (1990), and need not be placed in quotation marks. Williams and Bunkley-Williams (1990) also suggested that the term coral bleaching should be replaced by coral reef bleaching, since the events are not specific to corals, but affect a large group of photosymbiotic hosts that are an integral component of the coral reef ecosystem.
Reef Bleaching Global Patterns
According to Glynn (1991), coral reef bleaching events were reported as early as 1979-1980 from the Great Barrier Reef in Australia, the Ryukyus Islands in Japan, as well as from two Caribbean regions. However, these bleaching events were apparently very localized (i.e., island- or reef-specific), with the exception of the Ryukyus event, which was quite extensive. Without a doubt, it was the 1982-1983 circumtropical, El Niño-associated, coral reef mass bleaching event that stirred the scientific community into action. Bleaching was reported from: the Arabian region; the Mayotte and La Reunion Islands in the southwest Indian Ocean; Kepulauan Seribu and Karimunjawa Islands in the west and central Java Sea, respectively; the Ryukyus Islands in the northwest Pacific; the Tokelau Islands in the central Pacific; the Galapagos Islands and the Pacific coasts of Panama and Costa Rica; as well as in the Caribbean from Florida to Panama (Glynn 1991).
The biotic and abiotic effects of the 1982-1983 ENSO event, which was one of the most intense in recorded history, have been well documented (Barber and Chavez 1983; Halpern et al. 1983; Kerr 1983; Dandonneau 1986; Komar 1986; Glynn 1990). The flow of the warm western Pacific water into the cooler eastern tropical Pacific regions resulted in a significant sea surface temperature rise, that had immediate regional impacts in the eastern Pacific. The most severely impacted reef areas were in the Galapagos Islands (97% coral mortality) as well as along the Pacific coasts of Panama (75%-85% coral mortality) and Costa Rica (50% coral mortality) (Glynn et al. 1988). In all instances, coral mortality occurred during the peak period of high sea surface temperatures associated with the 1982-1983 ENSO event.
The 1986-1988 bleaching complex was even more extensive than the 19821983 event. Once again, bleaching occurred worldwide, but it was reported from a number of new sites extending from: Kenya in the western Indian Ocean; the Maldives Islands in the central Indian Ocean; the Andaman Islands in the Andaman Sea; Western Australia; Taiwan; and the Caribbean. The 1991-1992 ENSO event was associated with a bleaching event in the Society Islands, French Polynesia, where above-average temperatures caused bleaching in 53% of the coral genera, including 17% mortality. Throughout the 1980s, and early 1990s, coral reef mass bleaching events usually coincided with above-average sea surface temperatures, up to 2°-3°C above normal (Brown and Suharsono 1990), in many parts of the Indo-Pacific and the Caribbean (Glynn 1984, 1990, 1991; Fisk and Done 1985; Harriot 1985; Oliver 1985; Glynn et al. 1988; Jaap 1988; Goenaga et al. 1989; Williams and Bunkley-Williams 1990; Tsuchiya et al. 1992). Widespread coral mortality associated with bleaching is now a well documented, worldwide phenomenon (Glynn 1984, 1991; Brown 1987; Coffroth et al. 1990; Glynn 1990; Williams and Bunkley-Williams 1990) whose causative mechanisms are, however, very poorly understood.
It should be noted, however, that the apparent increase in reported bleaching events since the 1982-1983 El Niño event may be partially a result of greatly intensified efforts by the scientific community to document this phenomenon. Thus the 1986-1988 "coral reef bleaching complex", defined as a series of time-related bleaching events (sensu Williams and Bunkley-Williams 1990) associated with the 1986-1988 ENSO event, has been one of the best documented so far. Recent synthesis by Williams and Bunkley-Williams (1990) has shed new light on the geographic extent of this phenomenon. It is also clear that bleaching is not restricted to the scleractinian corals, but affects a number of other major coral reef phyla.
Reef Bleaching: Trigger Mechanisms
There are two schools of thought in the coral reef scientific community with regards to the primary causative factors for this phenomenon. Some scientists believe that the widespread, and more frequent, bleaching events are associated with global warming, while others maintain a more conservative view due to insufficient data. Williams and Bunkley-Williams (1990) have taken a strong position on the issue, and maintain that the global warming surge since the 1960s as well as significantly warmer conditions in the late 1980s, following the 1982-1983 ENSO event (Kerr 1988), were the primary cause of the 1980s bleaching complex "cycle". They point out that the two ENSO events of the 1980s (i.e., 1982-1983 and 19861988) provided atmospheric and oceanographic conditions that favored increased inshore temperatures above normal levels, which subsequently triggered the mass bleaching events. Based on their synthesis of a worldwide database of past bleaching events, Williams and Bunkley-Williams (1990) concluded that:
These conditions are not only well established, but seem almost certain to continue. The cycle may repeat in 1991 or 1992, possibly with more intensity, and will probably continue and increase until coral dominated reefs no longer exist.
Others accept that environmental stress is the main factor in the widespread bleaching events, but do not agree that bleaching related to abnormally high temperatures is associated with global warming. Atwood (1992) points out that "available data sets show no discernible warming trend that could cause such bleaching…". Note that the vast majority of bleaching reports are from coral reef areas where there is considerable concern with euthrophication and other anthropogenically-induced environmental stresses.
ZOOXANTHELLAE EXPULSION
The fact that corals respond to various environmental stresses by expelling their endosymbiotic zooxanthellae has been known for some time. How the zooxanthellae are expelled from coral tissue has been under investigation ever since the Great Barrier Reef Expedition in 1928-1929. These early studies suggested that, in corals exposed to stress (i.e., starvation and darkness), the zooxanthellae will first migrate through the endodermal tissue into the endoderm of the coelenteron, where they are subsequently released into the gastrovascular cavity and expelled to the outside environment through the mouth (Yonge and Nicholls 1931b). In the studies that followed, it became apparent that coral polyps were not only expelling zooxanthellae, but that they may actually be losing endodermal tissue (i.e., the host cells) along with their symbionts (fig. 11.12).
Recent studies have supported some of these earlier hypotheses, and demonstrated that bleaching in zooxanthellate sea anemones (Aiptasia pulchella) and corals (Pocillopora damicornis) is indeed associated with the expulsion of intact endodermal cells containing zooxanthellae, which degenerate subsequent to the expulsion (Muscatine et al. 1992). It was proposed that the expulsion associated with thermal stress is caused by dysfunction of the host cell adhesion mechanism (Muscatine et al. 1992). These findings are supported by studies done by Indonesian scientists who demonstrated that during bleaching, there is a loss of tissue organization and progressive reduction in the thickness of the endodermal layer. These studies suggest that tissue loss is either through expulsion or resorption. However, the most noticeable effect of bleaching is the gradual loss of zooxanthellae densities from the endodermal tissue (fig. 11.12C-D). The bleached tissue not only has a significant reduction in the abundance of host cells, but the cellular structure of zooxanthellae changes as well. Figure 11.13 clearly demonstrates that bleaching has a significant effect on zooxanthellae morphology.
In unbleached coral tissue (fig. 11.13A), the zooxanthellae in the endodermal layer exhibit a variety of morphologies, which are related to their different physiological states and natural mortality rates (Zd in fig. 11.13B) (Brown and Le Tissier, pers. comm.). During bleaching, the symbionts shrink considerably, which is reflected in large spaces in the vacuoles that harbour the zooxanthellae (fig. 11.13B). The shrinkage of zooxanthellae cells apparently results from an osmotic stress brought about by irradiance stress that upsets the natural balance between the host tissue and the zooxanthellae. Under normal conditions, the natural balance between the zooxanthellae and the host tissue is maintained by the transfer of photosynthates from the zooxanthellae to the host tissue (Brown and Le Tissier, pers. comm.). These observations are likely to be applicable to all bleaching events, since similar results have been reported from studies in the Caribbean (Hayes and Bush 1990). Note, however, that these studies may have been conducted in environments impacted upon by a variety of anthropogenic activities.

Figure 11.12. Induced bleaching in Acropora illustrating significant loss of zooxanthellae and endodermal tissue. A) Normal tissue with full complement of zooxanthellae in the endoderm. B) Temperature-induced stress results in expulsion of zooxanthellae. C) Loss of endodermal tissue and zooxanthellae is apparent. D) Significant reduction and breakdown of the endodermal layer. Ec ectoderm; M - mesoglea; En endoderm; Z - zooxanthellae. Scale bar 20 urn.
Photo courtesy of Suharsono, P30LIPI, Ancol, Jakarta.
Nature of Coral Bleaching
Whatever is the ultimate cause, or causes (i.e., stresses), of worldwide or local coral reef bleaching events, the main result is the expulsion of photosynthetic endosymbionts from the hosts and/or reduction of chlorophyll concentrations in the endosymbionts (e.g., zooxanthellae in corals). Bleaching is a stress response during which major changes in tissue organization and cytochemistry occur (Hayes and Goreau 1992). The final outcome of this process is a visible bleaching of the host (e.g., coral colonies become white), which can be either total or partial. It should, however, be noted that in some instances bleaching may be primarily related to the degradation of chlorophyll pigments in the zooxanthellae (and other photosymbionts), caused either by chloroplast disintegration or photooxidation of chlorophyll pigments (Asada and Takahashi 1987), rather than their expulsion. In corals, and many other photosymbiotic reef organisms, expulsion of the photosymbionts is a well documented stress response to elevated sea surface temperatures, or to thermal shock usually associated with a transient cold and heat stress (Mayor 1918; Jaap 1985; Steen and Muscatine 1987; Gladfelter 1988; Hoegh-Guldberg and Smith 1989; Kleppel et al. 1989; Brown and Suharsono 1990; Jokiel and Coles 1990; Szmant and Gassman 1990; Vicente 1990; Muscatine et al. 1992).
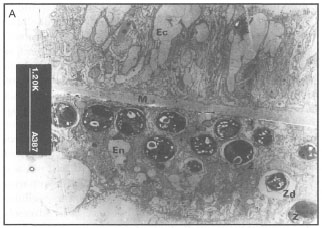

Figure 11.13. A) An unbleached coral tissue sample illustrating the general appearance of normal zooxanthellae. B) Bleached coral tissue illustrating zooxanthellae shrinkage as a result of osmotic imbalance. Note large spaces in the vacuoles harbouring the zooxanthellae. Ec - ectoderm; En - endoderm; M - mesoglea; Z - zooxanthellae; Zd - senescent zooxanthella; Vs - vacuole space.
Courtesy of B. Brown and M. Le Tissier, University of Newcastle, U.K.
The precise mechanism through which photosynthetic holobionts respond to environmental stress, which subsequently may induce mass expulsion of the photosynthetic endosymbionts, such as the zooxanthellae, remains largely unknown. There are a number of hypotheses that address this question, and these, as well as the nature of coral bleaching, have been recently reviewed by Brown and Ogden (1993). The current hypotheses may be viewed from either the coral's or the zooxanthellae's perspectives. It has been suggested that in response to environmental stress, the coral animal (i.e., the simple polyp) may evoke a number of physiological response (i.e., defenses) to maintain homeostasis. These responses (e.g., lower or higher metabolic rates), however, inadvertently alter the cellular micro-environment of the photosymbionts (e.g., nutrient and CO2 levels) to such an extent that the zooxanthellae will no longer find the association beneficial, and will as a result abandon their residence (Brown and Ogden 1993). In this scenario, bleaching is caused by a direct response of the photosymbionts to adverse cellular micro-environmental conditions, even though they may not have been adversely affected themselves by the original stress which, however, caused the coral animal to alter its metabolic function.
The alternate hypothesis is viewed from the animal's perspective. Under normal environmental conditions, the production of photosynthetically produced molecular oxygen, through zooxanthellae photosynthesis, often results in elevated PO2 in coral tissue. This seems to be a beneficial arrangement, since the coral polyp is sup plied with an abundance of oxygen for its metabolic needs. However, synergistic action of molecular oxygen with solar radiation (especially at the UV spectrum), mediated by photosensitizing agents, produces highly toxic forms of active oxygen (Lesser et al. 1990). The main toxic species of active oxygen are the superoxide radicals (O2-), hydrogen peroxide (H2O2) which results from the reduction of O2-, and finally, the hydroxyl radicals (HO•) (Lesser et al. 1990). These highly toxic substances have been shown to oxidize and denature various essential cellular compounds, such as lipids, proteins and nucleic acids, thus disrupting normal cellular function and eventually resulting in severe and irreparable cellular damage (Fridovich 1986). However, the zooxanthellae also produce specialized defensive enzymes (i.e., superoxide dismutase, catalase and ascorbate peroxidase) that act synergistically to deactivate both the superoxide radicals and the hydrogen peroxides, thus inhibiting the formation of hydroxyl radicals and preventing subsequent cellular damage (Lesser et al. 1990). Based on experimental work with a subtropical zoanthid, Palythoa caribaeorum, Lesser et al. (1990) demonstrated that increases in temperature and UV radiation significantly increase the activity of the defensive enzymes in the zooxanthellae. The observed increase in superoxide dismutase in the zooxanthellae is assumed to be an indirect indication of an increased production of active oxygen species. Since it is known that these toxic compounds can cross cellular membranes, it is likely that they are being exported from the zooxanthellae to the host cell (Lesser et al. 1990). Superoxide radicals and other species of active oxygen are very difficult to measure directly, therefore, we do not fully understand what may occur within the host cell. However, it seems likely that the subsequent expulsion of zooxanthellae from the coral tissue is a self-defense response by the coral polyp.
Based on available information it seems likely that bleaching is a result of a synergistic effect of a number of environmental parameters, notably temperature and irradiance (Lesser et al. 1990; Hagman Gittings 1992). In their experiments with Acropora valida and Pocillopora damicornis, Glynn et al. (1992) demonstrated that high seawater temperatures have a greater effect on the expulsion of zooxanthellae and the loss of chlorophyll and UV-absorbing pigments ("S-320") than high UV radiation. However, the study also demonstrated a significant interaction effect between high temperature and high UV radiation, indicating that sensitivity of corals to high UV radiation increases with increasing temperature, thus supporting the results of Lesser et al. (1990). Acropora valida was more sensitive to temperature rises than P. damicornis, a species known to inhabit shallow lagoonal waters susceptible to high daily temperature fluctuations. Interestingly enough, P. damicornis has been found to dominate shallow subtidal habitats on a recent lava flow (Gunung Api, Banda Islands) that is subjected to high-temperature (38°-42°C) plumes originating from numerous hydrothermal vents. Temperature sensitivity of some species may explain their absence from some regions influenced by the El Niño-Southern Oscillation events (Glynn et al. 1992), but their presence in other seemingly stressful thermal environments highlights the complexity of the temperature question.
UV Radiation
Corals, and other reef organisms, inhabiting shallow-water reef environments (e.g., intertidal) are exposed daily to high irradiance and ultraviolet radiation (UV), especially in the equatorial regions (Jokiel 1980). It has often been assumed that solar ultraviolet radiation does not penetrate much deeper than a few centimetres below sea surface, and thus the effects of UV radiation on deeper-water tropical communities were generally ignored, even though Jerlov (1950) demonstrated that, in fact, clear tropical oceanic waters allow considerable depth penetration (20 m) of UV radiation. This strictly optical property of seawater is a significant factor in the generally clear tropical oceanic waters. Furthermore, it is well-known that the equatorial regions have a relatively thin UV-absorbing ozone (a form of stratospheric oxygen) layer (Green et al. 1974; Baker et al. 1980), which is being severely influenced by massive anthropogenic emissions of chlorofluorocarbons (CFCs) into the atmosphere. Within the past two decades or so, Earth may have lost as much as 50% of its protective ozone cover, and the "ozone hole" over Antarctica has been estimated to be about half the size of Canada (McKay and Hengeveld 1990).
Of the total UV radiation reaching the planet, the earth's atmosphere (ozone and other gases) filters out effectively only the UV-C (200-280 nm) band. In the tropical regions, and under optimal environmental conditions, UV-A (320-400 nm) and UV-B (280-320 nm) radiation can penetrate through the water column and reach even the deeper (20-25 m) parts of the reef slope (Jerlov 1950; Smith and Baker 1979, 1981; Dunlap et al. 1988; Drollet et al. 1992). It is, however, unlikely that below 20 m depth UV radiation has much of a biological impact, since the water column at that depth is an effective filter. Gleason and Wellington (1992) have demonstrated that at a depth of 24 m, UV radiation has no effect on either zooxanthellae densities or chlorophyll concentrations, however, at depths less than 18 m, UV radiation may be an important factor. In their field experiments, Gleason and Wellington (1992) were able to show that zooxanthellae densities and chlorophyll concentrations were significantly reduced in corals exposed to ambient UV radiation when compared to UV-shaded corals. In a related study, Wellington and Gleason (1992) were able to show that at a latitude of 24.5° N (i.e., Bahamas), coral reef communities at a depth of 1 m can be exposed to maximum UV intensities (300-320 nm range) of about 1.4 W.m-2, while at a depth of 24 m the daily maximum intensities are reduced to 0.7 W.m-2. Note that these measurements were taken in a relatively oligotrophic environment, characterized by high seawater clarity. UV intensities in the equatorial regions, especially in the doldrums, may be higher. However, in more productive equatorial regions, especially along the extensive shallow-water continental margins, seawater transparency is considerably lower as a result of high concentrations of suspended particulate matter (SPM). Under these conditions, a significant portion of the incoming radiation may be filtered out, and therefore UV intensities, at least in the deeper parts of the reef, may be considerably lower. Unfortunately, comparable UV radiation data from Indonesia, or other highly productive 'upwelling' regions, are currently not available.
To cope with UV radiation, reef-building corals and other tropical shallow-water reef organisms (e.g., zoanthids -Palythoa tuberculosa) have evolved various biochemical defenses (Hirata et al. 1979). Specifically, it was discovered that corals are able to synthesize UV-absorbing substances, called S-320, which effectively filter out the UV-B radiation at a wavelength maximum of about 320 nm (Shibata 1969). Since this discovery, it has been shown that the S-320 substances offer corals a significant protection against UV radiation in the range from 290-340 nm (i.e., wavelengths responsible for skin melanoma in humans). The chemical nature of the S320 substances is being slowly unraveled, and we now know that the main ingredients of this efficient coral UV-filter complex are three mycosporine-like amino acids, namely, mycosporine-Gly (lmax310), palythine (lmax320) and palythinol (lmax332) (Dunlap et al. 1988) (fig. ll.14
Dunlap and Chalker (1986), working with a common Indo-Pacific coral, Acropora formosa, demonstrated that concentrations of S-320 in coral tissue were highest in shallow-water environments (1 m) and progressively declined with depth, with minimal concentrations at around 20 m. It seems that, at least for Acropora formosa, the two key S-320 compounds are the mycosporine-Gly (lmax310) and palythine (1 320). N /max
The UV-absorbing S-320 substances are an important protection, not only for the animal tissue, but also for the zooxanthellae, since UV radiation is known to cause photoinhibition and reduction in zooxanthellae growth (Jokiel and York 1984; Lesser and Shick 1989). It is a well-known fact that corals produce copious amount of mucus when subaerially exposed at low tides. Drollet et al. (1992) have recently demonstrated that coral mucus contains high concentrations of UV-absorbing compounds, especially within a few minutes of subaerial exposure, perhaps giving the polyps sufficient protection before they can retract deep within their carbonate skeletons for additional protection. However, bleached corals produce significantly less mucus than normal corals (Glynn 1990), and this may make them more vulnerable to the effects of UV radiation.
Lesser et al. (1990) found that increased temperature resulted in a significant reduction in the production of the UV-absorbing compounds, thus eventually leaving the holobiont more exposed to the harmful UV rays. Increased exposure to UV radiation will lead to even higher production of active oxygen compounds, thus intensifying the overall effect. Since exposure to higher UV radiation significantly decreases zooxanthellae production, Lesser et al. (1990) suggested that: "…reduction in the amount of translocated photosynthate decreases the benefit to maintaining the association." This seems to be a plausible explanation for partial bleaching, however, in cases where bleaching causes death of the host, this reasoning is harder to accept. In this respect, total bleaching and partial bleaching may be two loosely related phenomena, each with its own evolutionary significance.

Figure 11.14. The chemical structure of mycosporine-Gly, palythine and palythinol.
After Dunlap and Chalker 1988.
It is interesting to point out that coral UV-absorbing compounds may have found a prominent place in the world's sun protection industry, and it is more than likely that these powerful biochemical coral defenses will soon be protecting millions of humans along the shores of tropical seas (and elsewhere) from the Sun's cancer-causing UV rays. Mycosporine-Gly (lmax310) and palythine (lmax320) are among the most effective natural sun-screens found thus far, since their spectral coverage offers full protection against the most environmentally-harmful UV radiation in the 290-340 nm band.
Research into bioactive substances from tropical marine organisms is being vigorously pursued in many countries, even though rewards have so far been meager when compared to the wealth of new substances obtained from tropical rain forests. In Indonesia, basic research into marine-derived bioactive substances is still in its infancy, even though the economic value of biotechnology in future development is much talked about. For the time being, Indonesia, a nation with one of the highest marine biodiversity resources, and therefore, the highest potential for major discoveries, is a supply house for foreign research programs.
Bleaching in scleractinian corals, and other symbiotic organisms (e.g., Foraminiferida, Gorgonacea, Zoanthidae, Porifera and Mollusca), can be triggered by any number of natural environmental factors acting independently or, most likely, in synergy. Prolonged exposures to low or high light levels (Yonge and Nicholls 1931b; Dustan 1982), high UV radiation (Harriot 1985; Glynn et al. 1992; Hallock et al. 1992; Hallock and Talge 1993), abnormal salinities (Goreau 1964; Reimer 1971) and sedimentation (Bak 1978; Acevedo and Goenaga 1986) have all been known to cause zooxanthellae expulsion in corals. Disease cannot be ruled out as a contributing factor, since various pathogens have been isolated from bleached tissues of various symbiotic invertebrates (Glynn et al. 1985b; Goggin and Lester 1988; Jaap 1988). Extremely low tides and increased temperatures associated with ENSO events are also known to cause mass bleaching and extensive mortalities of intertidal reef organisms throughout the Pacific Ocean, especially in the western regions (Yamaguchi 1975; Glynn 1984; Tsuchiya et al. 1987). Lower-than-normal tides in the western Pacific during ENSO events are most likely associated with the easterly flow of the warm water pool that accumulates along the western boundary of the Pacific Ocean during non-ENSO years.
Coral Bleaching and Sea Level
Lower sea levels during ENSO events, as opposed to non-ENSO years, have been recorded in both the Timor and Flores Seas (Bray et al. 1994), but corresponding biological information is not available. Apparently, extremely low tides during the mass bleaching event of 1983 in the Java Sea (i.e., Kepulauan Seribu) have been ruled out. Harger (1994), however, points out that ENSO events (at least since 1976) seem to coincide with low sea levels in the Sunda Strait, and presumably in the Java Sea as well. Figure 11.15 demonstrates that there is a strong positive correlation between rainfall during the dry season in Java (i.e., June to October) and sea level.
The sea level in the Java Sea during the 1982-1983 ENSO event was about 10 to 15 cm below non-ENSO years. A 15 cm drop in sea level will expose a considerably greater area of the shallow inner reef flats that are characteristic of the Pulau Seribu coral reef system. Greater subaerial exposure potentially increases the local heating effect, especially during low tides, which will be translated into higher sea surface temperatures. Pulau Pari and Tikus, where the 1983 bleaching event was recorded, have a shallow lagoon that may have responded much faster to local weather conditions and experienced greater heating with a 15 cm drop in sea level. Higher temperatures, associated with calm weather, may have also significantly increased the evaporation rates during the day, thus creating daily pools of warm saline water that may have cascaded down the reef slope and affected corals in the deeper reef zones. One environmental factor that has not been taken into consideration is that the 1983 bleaching event in Pulau Seribu occurred during the period when low tides in the west Java Sea occurred during daylight hours (fig. 11.16).
It seems likely that the synergistic effect of above-average air temperatures, calm weather, above-normal evaporation rates and the timing of low tides during the hottest part of the day, may have significantly altered the normal hydrological conditions, thus triggering the bleaching response in corals and other reef symbionts.

Figure 11.15. Relationship between average rainfall during the dry period and sea level. Numbers correspond to ENSO year. Note that 1983 was among the strongest ENSO events on record.
Source: Harger 1994.
In an interesting bleaching study of the mushroom corals (Family Fungiidae) during the 1983 Kepulauan Seribu bleaching event, Hoeksema (1991) was able to demonstrate that fungiids inhabiting shallow-water environments (i.e., lagoons and inner reef flats) are less susceptible to bleaching than deeper-water conspecifics. During the Apriljune 1983 study, a period of maximum daily sea surface temperatures, no bleaching was observed at two lagoonal stations (< 4 m depth), and only 14% of fungiids were bleached on the inner reef flats (< 1 m depth). Goreau and Macfarlane (1990) found a similar spatial pattern of bleaching in Montastrea annularis in Jamaica, and suggested that shallow back-reef corals have a greater tolerance to high temperature and light exposure than deeper fore-reef corals. Figure 11.17, based on Hoeksema (1991), illustrates the percentage of fungiids (Fungiidae) affected by bleaching along a depth gradient around Pari Island.
The bleaching pattern demonstrated in figure 11.17 suggests that the cascading effect of warm saline waters may be a possible explanation. A similar phenomenon has been described from the Bahamas (Dennis and Wicklund 1993).
Oceanographic data collected during the bleaching event indicate that the above-average sea surface temperatures measured during March-June 1983 were restricted to the mid-late afternoon. Normal sea surface temperature values measured at 06:00 hours suggest that cooling may begin in late afternoon, and by nighttime sea surface temperatures over the reef may be close to normal values. Since temperature readings at 06:00 hours are not significantly different from the following non-ENSO years, or from previous years, it is likely that during March-June 1983 the region was influenced by unique atmospheric and oceanographic conditions that favored rapid heating of the shallow reefal waters during the day. Brown and Suharsono (1990) reported that the 1983 bleaching event was preceded by a few weeks of unusually cloud-free and calm weather, when maximum insolation is most probable. It is notable that bleaching was not observed in Pulau Seribu, or anywhere else in Indonesia, during the 1987-1988 and 1991-1992 bleaching events, which were otherwise global in proportions. Interestingly enough, there have been reports of wide-scale bleaching events (e.g., 1987-1988) that had no apparent environmental cause. However, Williams et al. (1987) did not rule out synergistic effects of temperature, light and ultraviolet radiation as possible triggers of these events.
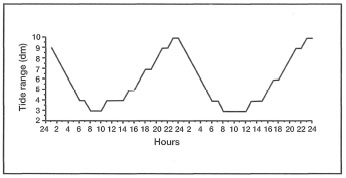
Figure 11.16. Hourly tidal fluctuations in Jakarta Bay and Kepulauan Seribu during May 1994. Low tides occur between noon and midnight from November to May, and from midnight to noon from May through October.
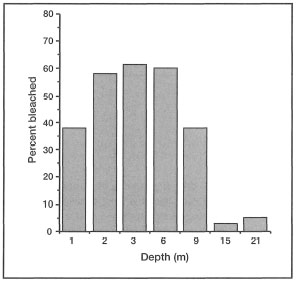
Figure 11.17. Percentage of fungiids affected by bleaching along a depth gradient. Sample size 2 x 10 m2 quadrats per sampling depth.
Source: Hoeksema 1991.
On a local (i.e., reefal) scale, occurrence of bleached coral colonies is a common phenomenon, even on undisturbed reefs and under normal environmental conditions. This type of bleaching seems to be more frequent in the intertidal and shallow-water habitats than on the deeper fore-reef habitats. The most frequently bleached species are small colonies of Acropora or Pocillopora. More puzzling is the fact that, even under normal environmental conditions, it is common to find two colonies of the same species, side by side, sometimes within a few centimetres of each other, with one colony fully bleached and the other with normal colouration. Many of the intertidal corals become exposed at low tides, and respond by retracting their tissue into their skeletons, which resembles bleaching, since the white carbonate skeletons become more exposed. Obviously this is a natural behavioral response to subaerial exposure (i.e., desiccation, exposure to higher temperature and UV radiation), and has nothing to do with loss of zooxanthellae.
However, the occurrence of bleached, or partially bleached, colonies in shallow-water subtidal reef habitats is more difficult to explain. Corals have a great capacity to maintain homeostasis under a wide range of environmental conditions, as is evident from their global and local distributional patterns. The association between the animal host and the photosynthetic symbiont must necessarily be dynamic in nature, where each partner must be tolerant to the other's physiological responses to different environmental stimuli. Bleaching may occur when this tolerance range is exceeded by a new (i.e., not previously expressed) response, by one or both bionts, to environmental conditions that are outside the normal range experienced by the holobiont. However, there seems to be substantial genetic variability (interspecific and intraspecific) that predisposes some groups, or individuals of the same group, to be more tolerant (i.e., having wider tolerance range) of certain environmental conditions than others. Frequent occurrence of bleached colonies in intertidal habitats may be a manifestation of this genetic variability. It has been shown that several coral species may have a number of genetically distinct strains, which may not be surprising, given the considerable phenotypic plasticity exhibited by a number of species groups. Earlier assumptions that different coral morphologies within given species are purely a response to environmental stimuli have been recently questioned (Tomascik 1990; Knowlton 1992). Perhaps not surprisingly, there is new evidence that the zooxanthellae symbionts themselves, as well as other endosymbionts, exhibit significant genetic diversity. Rowan and Powers (1991) have recently demonstrated that a number of Hawaiian scleractinian coral species contain distinctly different zooxanthellae, whereas the zooxanthellae of different colonies of the same species are the same. They also concluded that there seems to be greater zooxanthellae diversity between hosts than within hosts. This may explain some interspecific sensitivity, or predisposition, of certain coral species groups to bleaching, since different zooxanthellae strains may have different tolerance ranges to various environmental conditions. The intraspecific susceptibility to bleaching may, on the other hand, be a result of intraspecific genotypic variability, or a function of historical events and acclimatization to a specific set of environmental conditions.
During the 7th International Coral Reef Symposium, Buddemeier (1992) suggested that: "…sub-lethal bleaching is in fact an adaptive mechanism in that it permits the reinfection of the hosts with different species of algae that may confer superior stress resistance upon the symbiosis." An interesting bleaching event was observed in Barbados (Caribbean), on an offshore bank (1.5 km offshore) where, at a depth of 15 m, a large number of Dendrogyra cylindrus (on an average 5 to 10 m apart) became gradually bleached over a period of 12 days (Tomascik and Mah, unpublished). The bleaching started at the apex of each colony and progressed down to the base of the corallum. During this time, all other coral species exhibited normal colouration, and no evidence of bleaching or partial bleaching was observed. This event was clearly species-specific. All fully bleached colonies (some 1.5 m in height and 1 m in diameter) had their tentacles fully expanded and showed no "visible" signs of stress, such as low production of mucus (Glynn 1990) or tissue sloughing (Williams et al. 1987). Throughout the event, all bleached colonies responded to touch (i.e., diver's hand), by rapid withdrawal of all polyps. All environmental factors (depth 15 m), such as irradiance (PAR), seawater temperature, salinity, dissolved oxygen, pH and nutrients, were within normal range, with horizontal visibility at about 30 m. Histological examination of two ancillary tissue samples collected from two colonies (i.e., the apex of each colony) 12 days after the beginning of the bleaching event showed a total absence of zooxanthellae. About 26 days after the first sign of bleaching was observed, all Dendrogyra colonies were fully recovered in terms of their original colouration and without any mortality.
The selective nature of bleaching, as illustrated by the Barbados example, may not be as unique as it first appears. Tudhope et al. (1992) have recently documented an extensive bleaching event in Thailand, where seawater temperatures reached 2°C above normal, which is similar to the 1983 conditions reported in Kepulauan Seribu. During the bleaching event, a marked variation in the degree of bleaching of adjacent Pontes colonies was observed, suggesting that bleaching is selective (i.e., some colonies are more susceptible than others). Hoeksema (1991) also recorded bleached and unbleached individuals of the same species side by side during the 1983 Pulau Seribu event, and attributed this to intraspecific genotypic variability. The spotty nature of many bleaching events has been documented in other regions, however, the question as to why some colonies are more susceptible to bleaching than others has not been previously investigated. Tudhope et al. (1992) were able to demonstrate that bleached colonies exhibited slower skeletal extension rates (i.e., linear growth rates) than unbleached colonies. However, susceptibility to bleaching was not related to past growth characteristics (i.e., skeletal extension rates, skeletal density and calcification) of individual colonies., but is most likely related to other genetic differences. Goreau and Macfarlane (1990) also reported reduced growth rates and almost total cessation of calcification in Montastrea annularis following the 1987-1988 bleaching event in the Caribbean.
These examples of sub-lethal bleaching events seem to support the idea that in healthy corals, and perhaps in other symbionts, non-lethal expulsion of the photobionts may be a part of a natural process with an adaptive significance, as was suggested by Buddemeier (1992). Nonetheless, it is stressed that most bleaching studies conducted thus far (i.e., Kepulauan Seribu) are not undisturbed natural systems. A major shortcoming of all bleaching studies is the lack of environmental monitoring other than temperature. Sub-lethal bleaching may, therefore, be an acclimatization response by the individual to a new set of environmental conditions outside the range of earlier experience. The next bleaching event may not occur until the stress levels associated with the environmental conditions that triggered the acclimatization response are once again exceeded. The study by Hoeksema (1991) suggests that lagoonal and inner reef flat fungiids have been acclimatized to higher temperatures, and subsequently showed very limited bleaching. A similar example can be found in the Banda Islands, where a new coral community developed on Gunung Api's 1988 lava flow. During a 1990 visit, many of the coral colonies at a depth of less than 3 m were completely bleached, but alive (i.e., presence of mucus and visible tentacles). The maximum diameter of these colonies, which were mainly Acropora, Pocillopora and Pontes, was less than 10 cm. The most interesting feature in the area was the thermal regime influencing the coral community. Since the 1988 eruption, a number of shallow-water hydro thermal vents became active adjacent to the lava flow. Nutrient-rich warm-water plumes originating from these intertidal vents can periodically raise the ambient temperatures in the shallow-water subtidal areas (i.e., up to 1.5 m below the mean low water spring tide - MLWS) to 38° - 42°C. However, the lava flow coral community is also subjected to almost daily tidally-induced nutrient-rich upwelling events. During these events, water masses with temperatures at least 3°-4°C below ambient are spilled over the shallow reef. The overall structure of the thermal regime on the lava flow is one of high daily fluctuations. Thus, most corals in the shallow-water habitats on the lava flow will experience rapid and wide temperature swings on a daily basis. In a 1993 study of the same lava flow (Tomascik et al. 1996), the subtidal shallow-water habitat was covered by a profusion of coral colonies, with up to 60% coral cover. Most of the colonies present were again Acropora, Pocillopora and Pontes, which, however, became more abundant in the deeper parts of the lava flow. Among the large mature (up to 50 cm maximum diameter) unbleached Acropora, and smaller Pocillopora colonies, were numerous live colonies of bleached and partially bleached juveniles (fig. 11.18), while neighboring colonies (mostly acroporids) were fully pigmented.
However, coral bleaching was not observed (at comparable depths), on any of the adjacent and nearby fringing reefs sufficiently far away from the direct influence of the hydro thermal plumes. These observations suggest that sub-lethal bleaching, especially of juvenile corals, may indeed be part of a natural acclimatization process. Interestingly enough, Coles and Jokiel (1978) found that the highest survival rate of corals exposed to temperatures above 32°C were for those that were either partially or totally bleached during the acclimation period. This would suggest that bleaching may increase the thermal tolerance range of either the coral animal or the zooxanthellae, or both, by some unknown process. The common occurrence of partially bleached or fully bleached, yet healthy - from an anthropocentric viewpoint - colonies on Indonesian reefs may be a related phenomenon that awaits future studies.
One additional possible function of non-lethal bleaching that has so far not been investigated, relates to the reproduction of the zooxanthellae themselves. The question that needs to be addressed from an evolutionary standpoint, is 'what competitive advantage do the zooxanthellae obtain by the mass expulsions?'. We do not know whether the expelled zooxanthellae survive to reinfect another host colony, as is implied by Buddemeier (1992), or whether the event is in reality synonymous with "mass suicide" of the entire colony population. If the expelled zooxanthellae survive in seawater for any extended period of time, does sexual reproduction occur? Buddemeier's hypothesis suggests a major benefit for the host animal, but the benefits to the zooxanthellae are less clear. This is an exciting new area of research that needs immediate attention.

Figure 11.18. Bleached shallow-water corals on Gunung Api, Banda Islands. The site where this photograph was taken is within 1.5 m of a hydrothermal vent.
Photo by Tomas and Anmarie Tomascik.
Physiological Impacts of Bleaching
Since the symbiotic zooxanthellae play a significant role in a number of key metabolic functions (i.e., nutrition, waste removal and calcification) of the holobiont, it is expected that mass expulsion will have a negative impact on normal coral function. Cook et al. (1993) have recently suggested that zooxanthellae from partially bleached corals tend to be more nutrient sufficient than those from normal (i.e., unbleached) colonies. Nonetheless, some of the early physiological changes that have been documented to occur early in the bleaching event are increased metabolism (i.e., respiration rate), and decline in animal proteins, lipids and carbohydrates (Glynn et al. 1995a). Behavioral changes that occur are related to general tentacular lethargy and prolonged "uncharacteristic" tentacular withdrawal (Robinson 1985), however, Glynn (1990) found that polyps of all recently bleached coral colonies were expanded. An important physiological response to bleaching is a significant reduction of mucus (Prahl 1983; Glynn 1985b; Robinson 1985). Since mucus and lipid production in corals is linked directly with zooxanthellae carbon fixation rates and the subsequent translocation of a significant portion of the photosynthetically fixed carbon to the animal host (Crossland 1980; Crossland et al. 1980a,b), reduction in mucus and lipid production rates, following bleaching, should be expected.
Box 11.3. Coral bleaching in Indonesia.
Suharsono, P30-LIPI, Jakarta.
Animal-algal symbiosis is widespread among marine tropical invertebrates. About 150 genera of marine invertebrates, representing eight phyla, possess algal endosymbionts. The zooxanthellae (Symbiodinium microadriaticum) are yellow-brown unicellular symbiotic dinoflagellates which occupy the endodermal layer of all reef-building (i.e., hermatypic) corals as well as other reef-dwelling organisms. The relationship between the corals and the zooxanthellae is mutually beneficial.
The density of zooxanthellae in coral tissue appears to be relatively stable under constant environmental conditions. This apparent stability has been recently suggested to represent a dynamic rather than a static condition, reflecting a balance between growth rates of zooxanthellae and the host cells (Smith 1988). Regulation of this dynamic balance is thought to be determined primarily by nutrient levels, and available space in the host tissue.
Bleaching is a discoloration of reef-building corals which is caused by the loss of the endosymbiotic algae or zooxanthellae from the coral tissue. During bleaching, the colour of coral tissue changes from its natural colour (e.g., brown, green or blue) to bleached white or very light yellow, and is often followed by the death of the entire coral colony. However, under less stressful conditions, many bleached corals have been known to regain their zooxanthellae. Coral bleaching, or loss of zooxanthellae from coral tissue followed by colony death, has been associated with a number of natural environmental perturbations, such as prolonged emergence during extremely low tides (Loya 1976), increase in surface seawater temperatures above normal (Jaap 1979; Glynn 1983; Harriott 1985), or rain and associated salinity changes (Goreau 1964). Under natural conditions, bleaching is not necessarily followed by colony mortality, since many corals are capable of regaining their zooxanthellae and natural pigmentation. It has been demonstrated that scleractinian corals exposed to high temperatures for short durations are able to survive even if they expel all of their endosymbionts. However, prolonged exposures to high temperatures will usually result in permanent loss of zooxanthellae and the eventual death of the coral colony.
In general, most tropical marine invertebrates have a relatively restricted range of thermal tolerance, and are therefore considered as stenothermic. Elevation of temperature by as little as 3°-5°C above ambient has been shown to be detrimental to coral growth and survival in a number of tropical regions (Coles 1975; Coles et al. 1976; Jokiel and Coles 1977; Moore 1972). Ambient sea surface temperatures in the Indonesian Archipelago range between 25° to 31 °C, depending on the season, and provide optimal conditions for coral growth and reef development.
Indonesian archipelagic seas are relatively free from natural physical perturbations such as tropical cyclones, which can cause massive damage to coral reefs. However, between March and May 1983 a massive coral bleaching event associated with widespread coral mortality was observed throughout the Java Sea. The bleaching event occurred throughout an extensive area, stretching from the Sunda Strait (West Java) to Kepulauan Seribu (northwest of Jakarta) and the Karimunjawa Islands (Central Java) (fig. 11.19).
The first signs of bleaching were observed in March 1983 at Pari Island in Kepulauan Seribu. All bleached corals observed in March at Pari Island were still alive, since their polyps were extended normally and all colonies continued to produce mucus. In the early stages of the bleaching event the bleaching was very patchy, thus there were many instances when one colony of a particular species was fully bleached, yet a neighbouring colony of the same species appeared very healthy without any loss of zooxanthellae (fig. 11.20).

Fig 11.19. Map of the indonesianArchipelago. THe arrows indicate coral bleaching events in 1983.

Figure 11.20. The initial bleaching of coral colonies was observed in March 1983 at Pulau Seribu. Note the presence of healthy colonies adjacent to the bleached ones. As the event progressed, all colonies became bleached and a high percentage suffered mortality.

Figure 11.21. By the middle of May 1983, almost all branching corals were bleached. Acropora colonies suffered 80%-90% mortality.
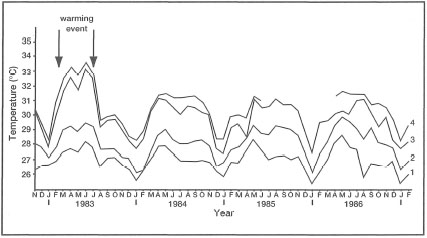
Figure 11.22. Average sea surface temperatures (°C) recorded at Pulau Seribu. Measurements were taken at: (1) 06:00; (2) 09:00; (3) 12:00, and (4) 16:00.
The first signs of a massive bleaching event were extensive patches of white or partially bleached colonies, extending from the reef flat down to a depth of 15 m. The most affected species were branching Acropora and Pocillopora, followed by hydrocorals (Millepora) and soft corals. By April 1983, approximately 40%-50% of corals on the reef flat were bleached, with a 10%-15% mortality. All dead corals were coated with a growth of turf algae and macro-algae.
Histological examination of the bleached coral tissue showed few, or an absence of, endosymbiotic dinoflagellates (i.e., zooxanthellae) in the endodermal layer, where they are very abundant under normal conditions. By the middle of May 1983, coral mortality on the reef reached 80%-90% (fig. 11.21).
By July, no further mortality was observed and some colonies started to recover. Pontes lutea and Montipora digitata were the first species to show signs of recovery, which was evident by the return of colouration to the tissue. The bleaching event affected 72 scleractinian coral species comprising 33 genera.
Regular monitoring of environmental conditions at Pari Island provided valuable information on sea surface temperatures during the bleaching event. The analysis of the temperature data revealed that the average sea surface temperatures at Pari Island were 2° to 3°C above average monthly values between March - August 1983 when compared to the same period in 1984, 1985 and 1986. Maximum sea surface temperatures at Pari Island were in excess of 33°C at mid-day and mid-afternoon (fig. 11.22).
Maximum daily sea surface temperature fluctuations ranging between 26.0°C to 31.5°C are common between May and August. The coral bleaching event in Pari Island and the subsequent mass mortality of scleractinians and other zooxanthellate reef organisms has been linked to the above average sea surface temperatures. The sea surface temperature records indicate that the warming event started about four to six weeks before the bleaching became noticeable. Since the bleaching event was widespread, from the reef crest to a depth of 15 m, and extended over a long period, the possibility of catastrophic low tide can be ruled out.
Szmant and Gassman (1990) have recently demonstrated that a significant number of corals fail to reproduce during the reproductive season following the bleaching event. Another response that has been reported is the resorption of coral tissue during prolonged bleaching events (Szmant and Gassman 1990). Figure 11.12 illustrates that tissue damage in bleached corals may be substantial, especially in the endodermal layer. Tissue repair, during and following bleaching events, requires major energy expenditure, which has to be diverted from other physiological functions that are less vital for the immediate survival of the individual colony (e.g., reproduction, growth, calcification). However, suppression of some of these functions may have significant long-term consequences with regards to community structure and competitive fitness. In long-term bleaching events, catabolism of endodermal cells can be considered as a sign of starvation, suggesting that the individual coral polyps may have either lost their ability to capture sufficient zooplankton or that zooplankton alone is not a sufficient dietary supplement for the loss of photosynthetically fixed carbon that is translocated by the zooxanthellae to the host animal. Alternatively, catabolism of damaged tissue may be more efficient than expending additional energy on food capture, at least during the "healing" period following bleaching events. Another problem that bleached corals must deal with is the removal of metabolic wastes, especially ammonia, since this metabolite is toxic at very low concentrations.
Coral Community Response
So far, we have reviewed the bleaching phenomenon from an organismal level as it mainly affects the holobiont complex, and may conclude that the potential physiological effects of bleaching are complex. Bleaching events, whether they result in mass mortalities of the affected fauna or not, cause significant changes in coral reef community structure and/or function (i.e., reef accretion and bioerosion). One of the first documented mass bleaching events in Indonesia occurred in Kepulauan Seribu during the 1982-83 ENSO event.
Coral community structure can also be affected indirectly by bleaching through the reduction or elimination of various coral invertebrate ectosymbionts, that "crustacean guard" (Glynn 1991). In a recent study, Tsuchiya et al. (1992) demonstrated that bleached corals not only loose their photosynthetic endosymbionts, but also their obligate crustacean ectosymbionts such as the xanthid crabs Trapezia and Tetralia. Their study supports results from earlier pioneering studies in the eastern Pacific region, which clearly demonstrated that increased predation by the corallivorous Acanthasterplanci was correlated with a significant decline of the obligate crustacean symbionts Trapezia spp. and Alpheus lottini (Glynn 1983, 1990).
Eakin (1993) showed that, following the 1982-83 ENSO event, the reefs along the Pacific coast of Panama were replaced by crustose coralline algae, which are much slower calcifiers (i.e., about 70% lower calcification rates than corals), thus reef accretion (i.e., growth) has been significantly reduced. Invertebrate bioeroders, such as sea urchins Diadema mexicanum, increased in abundance following the ENSO event and are having a negative impact on the reef structure, since their grazing activity has significantly accelerated erosion of the reef matrix. However, in many areas damselfishes protect the reef matrix within their algal patches, and reduce the erosion by as much as 70% (Eakin 1993). Similar conditions exist in Jakarta Bay, however, unlike in Panama, where rapid bioerosion is an ENSO-related phenomenon, bioerosion in Jakarta Bay is related to anthropogenic eutrophication (Tomascik et al. 1993).
There is a general public assumption that mass bleaching events are synonymous with mass mortality. In most instances, corals and coral communities will recover rapidly, provided they are not subjected to additional environmental stresses that may interfere with the recovery process. However, in some instances, mass bleaching events are followed by mass mortality of all bleached colonies (Glynn 1984). From the Indonesian experience, it seems that mass mortality events following mass bleaching seem to be restricted to relatively marginal environments, where the transient thermal stress is greatly intensified by existing environmental stresses, which can be either natural or, more frequently, anthropogenic in origin.
In the case of Kepulauan Seribu, coral mortality and the subsequent slow recovery of the shallow-water coral communities is most likely related to the degradation of environmental conditions associated with massive development in the area as well as increased pollution from Jakarta. It seems that the 1983 mass bleaching event was restricted to the Java Sea, since no reports of the same phenomenon were received from any of the major diving destinations in Indonesia (e.g., Manado, Banda Islands, Bali, etc.). Review of sea surface temperature data during the same period in eastern Indonesia indicated that normal temperature conditions prevailed in areas monitored at the time.
The above discussion relates mainly to temperature as the leading factor in most mass bleaching events. More insidious, however, are anthropogenic activities that result in high sedimentation rates, high turbidity, excess nutrients, low salinities and oxygen levels, all characteristic of well-documented (Brown and Howard 1985) impacts of industrial, agricultural and urban pollution. There is no question that these anthropogenic activities significantly impact upon coastal reef ecosystems (e.g., Jakarta Bay) (Tomascik et al. 1993) and that they may be a major factor in either initiating local bleaching events, or significantly interfering with recovery from naturally induced effects. The well-documented example from Kepulauan Seribu (Brown and Suharsono 1990; Warwick et al. 1990) clearly suggests that anthropogenic influences cannot be dismissed, or equated with normal environmental perturbations (Grigg and Dollar 1991). Nevertheless, there seems to be strong evidence that widespread "regional" coral bleaching events may be directly linked to abnormally high sea surface temperatures, whose causes are, however, not fully understood (Glynn 1990, 1991; Glynn and D'Croz 1990; Glynn et al. 1992; Hagman and Gittings 1992).
VOLCANISM, EARTHQUAKES AND TSUNAMIS
Volcanism
Sumatra, Java, Bali and Lombo;, four names that immediately conjure up images of high volcanic cones towering over extensive paddy fields (fig. 11.23). Indeed, these islands are world famous for some of the most breathtaking scenery, which is a mixture of the sublime and tranquil beauty of the deep-green paddy fields, and the awesome power of the volcanoes that are looming ominously, with a strange eerie peacefulness, in the background. Throughout the vast archipelago, the dominant role of the volcanoes is clearly evident through the cultural fabric of many island people (e.g., Balinese). This is perhaps not surprising, considering the fact that the inhabitants of this seeming tropical paradise have for centuries been subjected to the devastating power of volcanic eruptions, which have occurred frequently and unpredictably in the past (table 11.1).
One of the largest volcanic eruptions in the archipelago occurred sometime in the Pleistocene (100-75 ka B.P.), and resulted in the formation of Lake Toba, which today is an important tourist destination in North Sumatra. Lake Toba is an old caldera formed by a massive volcanic eruption which released between 1000-2000 km3 of ejectamenta. The impact of this eruption on the region and its biota can only be guessed at. More recently, the 1815 eruption of Tambora (Sumbawa) released between 150-300 km of ejectamenta (total energy release of about 8.4 x 1026 ergs) causing a major cataclysm which resulted in 92,000 casualties. Hundreds of kilometres to the southwest, the deep Indian Ocean sediments have faithfully recorded the wrath of Tambora. And how can one forget the well-known, and much-written-about, Krakatau eruption of 1883 in the Sunda Strait, which released about 18 km of ejectamenta with a force of 125 megatons of TNT (i.e., about 1 x 1025ergs), even though it was the smallest of the three (fig. 11.24). Krakatau even made it to Hollywood, starring in the film Krahatoa - East of Java, although geographical accuracy obviously was not allowed to interfere with artistic license.

Figure 11.23. Fields of deep-green paddy and rich tropical vegetation hide much of Bali's volcanic foundations, while only clouds shroud Gunung Agung.
Photo by Tomas and Anmarie Tomascik.
GEOGRAPHIC DISTRIBUTION OF VOLCANOES
The Indonesian Archipelago is a tectonically active region, and thus contains many islands with active volcanoes. There are over 500 known volcanoes in Indonesia, of which 80 are considered as active and 50 are in the solfataric and fumarole stage (van Bemmelen 1949; Hutchison 1989).

Figure 11.24. The 1993 eruption of Anak Krakatau in Sunda Strait.
Courtesy of A. Herwanto, Puslitbang Geotechnology LIPI, Bandung.
Table 11.1. Regions and areas that are under potential threat of volcanic eruptions.
Table 11.2 lists all the major active volcanoes in the archipelago with recorded eruptions since 1900. Of special interest are the oceanic island and submarine volcanoes of the eastern (e.g., Inner Banda Arc - Gunung Api, Banda Api, Serua, Nila, Manuk), and northern (e.g., Gulf of Tomini - Una Una; Sangihe Islands - Awu, Siau, Ruang, Banua Wuhu) regions of the archipelago, since their activity has a profound effect on the life-death cycle of coral reefs and coral communities that surround them (figs. 11.25 and 11.26).
Volcanism and Coral Reefs
It has often been suggested that active volcanism generally inhibits coral reef development (Davis 1928), and this certainly seems to be true in many regions, since the development of coral reefs on volcanic islands is under the influence of a variety of abiotic and biotic factors that vary across spatial and temporal lines.
… on the coasts of volcanic islands conditions must be adverse to the growth of corals, on account of the large quantities of erosion products that are continually transported to the sea. I saw a very convincing proof of this thesis at Krakatoa.—UMBGROVE 1947
However, when viewed from a broader regional perspective, volcanism (i.e., eruptions and associated lava flows) may not necessarily be detrimental to scleractinian coral communities, even though the development of the coral reef structure (e.g., the fringing reef) may locally be set back considerably. Under appropriate environmental conditions, scleractinian coral communities are known to exist and persist without a coral reef structure, provided a suitable substrate (not necessarily solid) is present - and lavas may be such a substrate. The impact of volcanism on coral reefs and coral reef-associated communities is obviously a matter of magnitude and frequency of eruptions as well as seismic activity.
Table 11.2. Active volcanoes of the Indonesian Archipelago. Table contains only volcanoes with recorded eruptions since 1900 (i.e., active solfatara and fumarole volcanoes are not included). 'Year' refers to the last recorded eruption.

Figure 11.25. Two active volcanoes from the northern and southern regions of the Indonesian Archipelago. A) Gunung Siau, Sangihe Islands; B) Gunung Lewotolo, Lembata Island, East Nusa Tenggara.
Photos by Tomas and Anmarie Tomascik.


Figure 11.26. Many volcanoes have a direct impact on the surrounding coral reef communities and other coastal ecosystems. A) The 1988 lava flow on the northern slope of Gunung Api (Banda Api), Banda Islands. B) The 1949 lava flow on the northeast slope of Gunung Ruang.
Photos by Tomas and Anmarie Tomascik.
Figure 11.27. The history of Krakatau volcano (looking from north to south). 1A) The hypothetical original andesitic volcano; 1B) Early caldera collapse forming the base of the present islands; 2) The growth of smaller basaltic Rakata volcano; 3A) The formation of Krakatau from merging of Rakata, Danan and Perbuwatan volcanoes.; 3B) Krakatau after the August 26-28, 1883 eruption; 4) Formation of "Anak" Krakatau, which emerged December 1927.
From van Bemmelen 1949, p. 201.
The 1883 Plinian eruption of Krakatau is a good example of a major local environmental perturbation that eliminated all coral reefs that surrounded the island. Krakatau (also known as 'Krakatoa') actually consisted of three volcanoes (i.e., basaltic Rakata and andesitic Danan and Perboewatan) prior to the eruption (fig. 11-27).
The eruption caused rapid evacuation of the magma chamber, with subsequent collapse of the volcanic complex into its empty chamber. Thus, Krakatau Island proper was not really blown apart, as is often assumed, but rather, it rapidly subsided below sea level with the exception of a small section of Rakata Island. According to Sluiter (1890), scleractinian corals (Acropora spp., Pontes spp. and Favia spp.) started to colonize an exposed basaltic lava slope along the northwest coast of Rakata within five years of the eruption. However, a 1929 resurvey of the same site by Umbgrove (1947) showed that none of the corals recorded by Sluiter in 1989 were present, and that the area in general was devoid of reef growth and corals. Umbgrove (1947) attributed this to the unfavorable environmental conditions and unstable substrate.
One hundred years later, reef development in the former Krakatau area is marginal. Nonetheless, about 213 scleractinian species have been recorded from the various islands in Sunda Strait and Lampung Bay, South Sumatra, but only one location was sampled in the Krakatau Islands southwest of Pulau Sertung (Sukarno and Suharsono 1983).
An earlier survey of the Krakatau Islands in 1981 provided a qualitative estimate of coral species richness at 10 widely spaced locations, with a total search time of 390 minutes (Salm et al. 1982). The results of the two surveys are very different and are presented in table 11.3 ('Scleractinian coral species of Krakatau Islands') to demonstrate the need for standard sampling protocol and greater taxonomic refinement. While the list is not very useful for quantitative comparisons with other regions, it may be used, with great caution, to compare the general community structure with other volcanic islands in Indonesia. Based on these surveys, it seems that Krakatau, as a region, has a relatively diverse assemblage of scleractinian corals belonging to 62 genera represented by 97 species (Salm et al. 1982). However, at a local (i.e., reef site) scale the reefs are generally impoverished. The average species richness per location was only 48.6 (STD±9.2) scleractinian species, while the average generic richness was 33.4 (STD±7.4) genera. This pattern seems to reflect high habitat heterogeneity within the area (100 km2). While the overall diversity seems to be high, considerable sampling effort (in time and space) was used to obtain the results. Note that in the 1981 study, Acropora, Montipora, Goniopora, Favites, and Goniastrea were not identified to species level, because of taxonomic complexity of these groups, even though they are among the most species-rich coral genera. Thus, the high generic diversity, when compared with species richness, is an artifact of sampling, and is not related to any unique environmental conditions.
The lack of taxonomic detail of earlier studies conducted in Indonesia, with a few exceptions, is unfortunate, because regional quantitative comparisons (biogeographical or ecological) are difficult, even though a great wealth of data seems to be available. For example, the 1981-1982 Krakatau results (Salm et al. 1982) cannot be meaningfully compared with the results of the 1993 Banda Api lava flow study, mainly because of different sampling techniques (i.e., qualitative vs. quantitative), effort (large areas surveyed vs. restricted to limited number of line transects), area of the island group (100 km2 vs. 25 km2), and taxonomic detail (trainees vs. coral taxonomist). Using qualitative sampling techniques similar to those employed by Salm et al. (1982) in the Banda Islands would significantly increase both generic and species diversity.
With the exception of the 1981 and 1983 studies, not much is known (quantitatively or qualitatively) about the current status of Krakatau scleractinian coral communities, especially after the recent activity of Anak Krakatau that has already claimed a number of lives. Based on our qualitative observations in 1992 (Tomascik and Mah, pers. obs.), the scleractinian diversity along the southwest coast of Rakata Kecil (Pulau Panjang) is relatively high, considering the marginal conditions of the environment. However, both earlier studies have greatly underestimated the diversity of both Acropora and Montipora in the area.
Water-quality conditions in the immediate vicinity of Anak Krakatau seem to be marginal, mainly due to the erosion of tephra deposits as well as leaching of iron oxides from Anak Krakatau's lava flows, which frequently colour the water muddy brown. Furthermore, in contrast to the volcanoes in the eastern and northern regions of the archipelago (Banda Api and Ruang) which release andesitic lava (i.e., viscous, acidic and high SiOs content), the lava of Rakata has been classified as basaltic (i.e., low viscosity, basic and low Si02 content).
Based on our visits to a number of active volcanoes (subaerial and submarine) in the northern and eastern regions of the archipelago, it seems that the relatively depauperate coral fauna of the Krakatau area is an exception, rather than a rule. All other active volcanoes visited (Gunung Api, Nila, Serua, Damar, Ruang, Siau, Mahengetang and Manuk) showed a remarkable coral species diversity that was associated with high coral cover, which in places surpassed 100% as a result of canopy-like growth. Based on these observations, it seems probable that the steep lava slopes of andesitic Plio-Pleistocene volcanoes may have served as important refuges for scleractinian corals during the widely fluctuating sea levels, and today these volcanic islands support some of the most diverse hermatypic coral reef assemblages in the world. Table 11.4 lists the major submarine centres of volcanic activity in the archipelago.
Banda Api: Case Study
The Gn. Api (Banda Api) is one of the most active volcanoes of the Dutch East Indies. …the reefs between Banda Naira and Lonthor [Banda Besar] are normal flourishing reefs which show no signs of being troubled by their very close proximity to the Banda volcano. It is not known whether reefs grow on the volcano itself. …As long as the reefs are not situated on the direct slope they can exist quite close to a volcano that has seen many recent ash-eruptions.—KUENEN 1933
The fabled "Spice Islands" of the East Indies were the original source of precious nutmeg and clove, whose trade had a significant impact on European history. The nutmeg tree, Myristica fragrans, requires special conditions which, from an evolutionary point of view, must have been optimal in these islands. One of the primary requirements of high-quality nutmeg is rich volcanic soil, for which there is no shortage on the Banda Islands, since the entire mini-archipelago is of volcanic origin. The Banda Islands consist of six islands, namely, Banda Naira, Banda Besar, Gunung Api, Hatta, Ai and Run, with an approximate land area of 8150 ha. The nutmeg tree still flourishes on the islands; currently there are about 150,000 nutmeg trees on the Banda Islands alone, but the remoteness of the islands and the establishment of large nutmeg plantations on Java and other islands of the archipelago, closer to the market, has considerably reduced their earlier economic significance. Even though Indonesia supplies 75% of the world's nutmeg, with Grenada supplying the balance, the importance of the Banda Islands has declined, but not the quality of the nutmeg spice, which many consider the best in the world.
The Banda Islands, because of their remote location, are among the very few areas in the world where the marine and coastal resources, primarily coral reefs and seagrass beds, are still in a pristine condition. The main reason for the unspoiled nature and beauty of the islands, in addition to geographic isolation, is that during most of the Bandanese history the inhabitants did not rely on the sea (i.e., the coastal resources) for survival, but rather were supported by the flourishing nutmeg trade. As a result of the unspoiled nature of the marine environment, the Banda Islands are a world-class diving destination that offers breathtaking underwater scenery on some of the most diverse coral reefs in the region.
Table 11.3. Scleractinian coral species of Krakatau Islands. A: Results from a 1981 study of 10 locations in the Krakatau Islands using qualitative assessment techniques. B: Results from a 1982 study at Pulau Sertung (Krakatau Islands) using quantitative survey using 10 m line transects.


Table 11.4. Major submarine centres of volcanic activity in the Indonesian Archipelago. Depth indicates the approximate depth of the volcanic summit.
The "Spice Islands" are beginning to attract considerable new interest from the coral reef scientific community as a result of a volcanic eruption that occurred in May 1988. The Banda Islands (04°30'00" S 129°53'00" E) are located in the eastern Banda Sea on the inner volcanic Banda Arc that separates the deep Banda and Weber Basins. At the centre of the island chain is an active volcano named Gunung Api (Banda Api), or the 'Fire Mountain' (not to be confused with Gunung Api located north of Wetar) (fig. 11.28).
Gunung Api, or Banda Api, has an elevation of about 656 m above sea level, and is considered to be one of the most active volcanoes in eastern Indonesia (van Bemmelen 1949). Together with the neighboring islands of Banda Naira and Banda Besar, Gunung Api is an active remnant of a former larger volcano (Kuenen 1933a). After being dormant for nearly 97 years, Gunung Api suddenly became active on April 20, 1988, and erupted on May 9, 1988 (Casadevall et al. 1989; Pardyanto et al. 1991).
Gunung Api has a long history of violent eruptions that occurred in 1891, 1810,1720 and 1614 (Kusumadinata 1979). The 1988 eruption commenced with a phreatic eruption at the summit crater, with an ejection of thick white smoke to a height of about 200 m above the crater (Pardyanto et al. 1991). Throughout the day a series of explosive eruptions, Plinian in behaviour, occurred from five vents that released approximately 8 x 10 m of lava during the first nine hours of the eruption. The Plinian eruption column rose to an altitude of 16,000 m (Casadevall et al. 1989). The main eruption lasted less than 12 hours, but was followed by 10 days of strombolian explosions mainly from the summit crater (Pardyanto et al. 1991).
Fortunately for the inhabitants of Banda Naira, the main eruption occurred on the southwestern slope of the volcano. The vegetation on the western slope of the volcano, however, suffered total decimation, while the eastern part was not affected, mainly as a result of strong easterly winds. The marginal fringing reef along the exposed southwest coast was smothered by large quantities of pyroclastic deposits (i.e., tuffs and volcanic ash deposits). The extent of heat-associated mortality during the eruption is not known, however, clear evidence of thermally-induced deaths of shallow-water (i.e., 0-2 m depth) corals as well as partial mortality of corals on deeper slopes (depths of 2-4 m) were observed along the northwest coast of Banda Besar, over 1 km across the Sempitan Lonthor (i.e., Lonthor Passage) (fig. 11.29).
Since much of the southwestern coast is fully exposed to swell and rough seas generated during the Southeast Monsoon (April - November), rapid erosion occurred soon after the eruption, resulting in submarine topography characterized by an aggregate of unstable lava rock, large lava boulders and ash.
The eruption of Gunung Api released large quantities of viscous (acidic) andesitic lava (Casadevall et all. 1989; van Bergen et al. 1989; Pardyanto et al. 1991). The largest lava flow, with a volume of about 3.2 x 106 m3, originated from a north vent at an altitude of 200 m, and entered the sea at 11:00 on May 9, 1988 (Casadevall et al. 1989). The flow rate of the northern lava was estimated at 0.4 x 106 m.h-1 and continued for several days (Casadevall et al. 1989). The north lava flow was fed through a tube system, and upon entering the sea it covered about 40,000 m2 of a well-developed fringing reef (Sutarna 1990). Because of the local topography, the lava flow split into two branches just before reaching the beach, thus leaving an approximately 150-m-wide "island" consisting of the original reef matrix (fig. 11.30). Since this part of the coastline is protected by a headland and a small island from seas generated during the Northwest and Southeast Monsoons (i.e., December - March and April - November, respectively), no visible erosion or break-up of the lava flow occurred during the five years following 1988. The resistance of andesitic lavas to weathering and erosion may be a key factor in facilitating rapid colonization, since without erosion there is virtually no sedimentation or siltation that can retard coral recruitment, and the subsequent reef growth. Gunung Api is not an isolated case, since similar conditions occur at Ruang Island (Sangihe Archipelago) where a similar eruption occurred in 1949, yet the lava remains intact without any visible signs of major weathering or erosion (fig. 11.31).
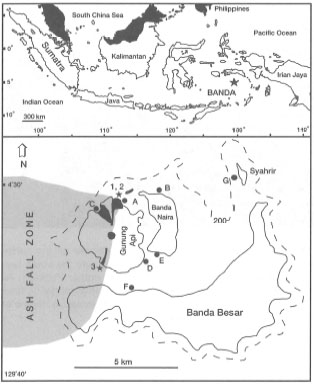
Figure 11.28. Map of the Indonesian Archipelago showing the location of the Banda Islands (upper), and the location of the eruption (lower). Locations numbered 1 to 3 (H) were surveyed in this study, May 1993. Sites labeled 'A' to 'G' (I) were surveyed in November 1986 (Sutarna 1990). Areas shaded black represent the approximate location of the crater and the northern parasitic cone. Dotted line represents the 200 m depth contour.
The natural calamity at Gunung Api provided a unique opportunity to conduct a baseline study that will serve as an important base for a long-term study to monitor the development of this new coral community and the embryonic coral reef. In collaboration with Dr. Robert van Woesik and Sandy van Woesik we embarked on an expedition to study the 1988 lava flow at Banda Api, which proved to be a fascinating site.

Fjgure 11.29. Massive colony of Diploastrea heliopora on the northwest coast of Banda Besar (Lonthor), about 1 km from the south lava vent on Gunung Api (see fig. 11.25) across the Lonthor Passage. All massive colonies between 2-4 m depth suffered partial mortality, from which a majority partially recovered by 1993. All colonies at depths of less than 2 m suffered total mortality.
Photo by Tomas and Anmarie Tomascik.

Figure 11.30. Gunung Api, Banda Islands. Aerial view of the 1988 lava flow on the northwest slope of the volcano, May 1993. Note the non-erosional lava coastline.
Photo courtesy of R. van Woesik, University of Ryukyus.

Figure 11.31. An andesitic lava flow which has remained virtually unchanged (i.e., limited erosion) since it entered the sea following the 1949 eruption at Ruang Island, Sangihe Archipelago, North Sulawesi.
Photo by Tomas and Anmarie Tomascik.
Coral Colonization of the 1988 Lava Flow. Earlier estimates of coral community recovery from various environmental disturbances of different intensities range from 10 to 70 years (Pearson 1981; Colgan 1987; Done et al. 1991; Dollar and Tribble 1993). However, disturbance and recovery of scleractinian coral communities following volcanic eruptions have received only scant attention (Grigg and Maragos 1974), yet volcanism has been recognized as a primary geological process that facilitated the dispersal of coral reefs throughout the tropics by creating new substrates (i.e., islands or 'stepping stones') for coral colonization (Darwin 1842; Dana 1872; Davis 1928). On the other hand, volcanic eruptions and lava flows on volcanic islands surrounded by coral communities are also major physical disturbances, since lava flows and volcanic ash may cover entire reef tracts and destroy all benthic life in their path. Volcanic eruptions, especially the lava flows associated with them, are a unique disturbance, since apart from large-scale disturbances that are measured on a geological time scale (e.g., glaciation), other environmental disturbances, such as tropical cyclones (Woodley et al. 1981; Mah and Stearn 1986; van Woesik et al. 1991), short-term sea-level anomalies (Loya 1976), El Niño events (Glynn 1984,1990; Glynn and Colgan 1992), population explosions of coral predators (e.g., Acanthasterplanci - Done 1985; Faure 1989; Done et al. 1991; Drupella cornus - Moyer et al. 1982) or mass mortality of invertebrate herbivores (Diadema antillarum) (Lessios et al. 1984), do not destroy 100% of all benthic species within the impact zone, nor significantly alter the chemical composition of the reef matrix.
However, not all volcanic eruptions are associated with lava flows, which may significantly enhance coral recovery by providing new substrate for subsequent colonization. For example, Pajaro et al. (1993) reported that the eruption of Mt. Pinatubo had a significant negative impact on the benthic coral communities at Zambales, Philippines. Live percent coral cover following the eruption declined from 47.7% to 9.9% at a depth of 3 m, while at a depth of 10 m coral cover decreased from 50.8% to 9.7%. A considerable part of the reef was buried by ash deposits, thus increasing abiotic cover from 43.8% to 87.7%. Reefs closest to the eruption were the most adversely affected. Note that lava flows reaching the coastline were not reported. In contrast, coral reef communities at the Banda Islands during the 1988 eruption of Gunung Api did not suffer widespread damage. However, there was a noticeable reduction in coral cover along the southwest coast where the fringing reef was buried by thick deposits of tephra and loose lava rock. Casadevall et al. (1989) reported that the south side of Gunung Api was covered at places by up to 36 cm of tephra, but reefs on the adjacent islands downwind showed no evidence of negative impact, since most of the ejected tephra fell into open ocean.
In a 1986 study conducted in the pre-eruption Banda Islands, Sutarna (1990) recorded a total of 86 scleractinian coral species from seven locations (table 11.5.) His results are in contrast to our study conducted in 1993, where 138 Coral species were recorded at the three locations situated on Gunung Api. By coincidence, one of the Sutarna's 1986 study sites (i.e., site A-fig. 11.28) was apparently located within 300 m of the new lava community and the old reef that was not covered by the lava flow. The 41 coral species recorded at Sutarna's site in 1986 may be considered as roughly representative of the coral species richness that existed in this particular area before the 1988 volcanic eruption (Sutarna 1990). However, the data need to be interpreted cautiously, since there are differences between the two studies with regards to the level of replication and sampling effort. However, a general qualitative comparison is instructional, and the results clearly indicate that while the lava flow destroyed the original reef (which may not have been that old considering the frequency of Banda Api's eruptions), it created a local, if not regional, hot spot of coral diversity.
The coral community at location A in 1986 was dominated by massive forms of Pontes lutea and Diploastrea heliopora. Scleractinian corals accounted for about 37% of the total substrate cover (24% by massive forms alone), while soft corals accounted for 19% (Sutarna 1990). These findings are in contrast to our results for the lava flow, where coral species richness (i.e., 124 species) and coral coverage (61%) have increased. In contrast to the rapid coral community development that occurred on the lava flow, recovery of the scleractinian coral community on the adjacent old carbonate matrix has been much slower. While species richness on the old carbonate reef (45 coral species) has reached comparable levels recorded at location A in 1986, the scleractinian coral coverage (8.3%) was well below the 1986 value (37%), and soft corals were almost totally absent from the community.
The 10 locations investigated during the 1986 and 1993 surveys split into two groups reflecting significant habitat differences (fig. 11.32). These differences are mainly related to the degree of exposure to the sea. For example, cluster "a" groups coral communities from sheltered sites that, in terms of hydrological conditions, are characteristic of well-protected lagoonal environments. While the coral communities are protected from heavy seas throughout the year, they are nonetheless subjected to relatively strong tidal currents (50 to 250 cm/sec) (Tomascik and Mah, pers. obs.). Coral communities that developed in these relatively stable lagoonal environments may represent climax communities that, however, differ from one another in species composition at comparatively short distances (10's of m). The extensive coral communities off the southwest coast of Banda Naira form an intricate mosaic of monospecific stands with 100% cover - or more. The boundaries between the various monospecific stands are knife-edge sharp, yet no apparent environmental gradient can be seen. What environmental or biotic factors cause this peculiar distribution in this area is not known. The dominant species are Acropora formosa, A. subglabra, A. carduus and Pavona cactus. The common features of the environment are the calmness of the sea surface and fast-flowing tidal currents. As would be expected, coral communities characteristic of more open and exposed environments form a distinct group (i.e., cluster "b") of their own.
Figure 11.32. Dendrogram showing classification of the 1993 and 1986 study sites by the Ward minimum variance method (SYSTAT), using normalized Euclidean distance (root mean squared distance) as the measure of dissimilarity between sites (Wilkinson 1989). Community descriptors used to characterize each site were: 1) the total number of species; 2) percent cover by Acropora spp.; 3) percent cover by branching corals other than Acropora spp.; 4) percent cover by massive corals including Porites spp.; and 5) percent cover by foliose corals. Site designation as for fig. 11.28.
The 1993 quantitative study of the Banda Api lava flow demonstrated that the coral community that became established on the northern lava flow had a significantly higher average number of coral species, percent coral cover, colony abundance and coral diversity than either the adjacent reef substrate that was not covered by the lava, or the unstable tephra deposits on the southwest (high-energy) coast of the volcano (figs. 11.33, 11.34, 11.35 and 11.36).
It is often stated that acroporids are highly successful opportunistic species, and this is the case on the Banda Api lava flow. Acropora spp., and other branching species, were the most successful colonizers on both the lava flow and the old carbonate reef matrix with respect to colony abundance (fig. 11.37). However, species with encrusting morphologies were more abundant at the old carbonate reef than on the lava flow.
Table 11.5. Qualitative distribution of scleractinian and non-scleractinian corals in the Banda Islands based on presence (+) and absence (-) data from Gunung Api, Banda Naira, Banda Besar and Syahrir. Data for locations 1 to 3 from May 1993, five years after the eruption of Gunung Api. Data for locations A to G from November 1986, approximately 1.5 years before the eruption of Gunung Api (Sutarna 1990). Sampling site locations as for figure 11.28.
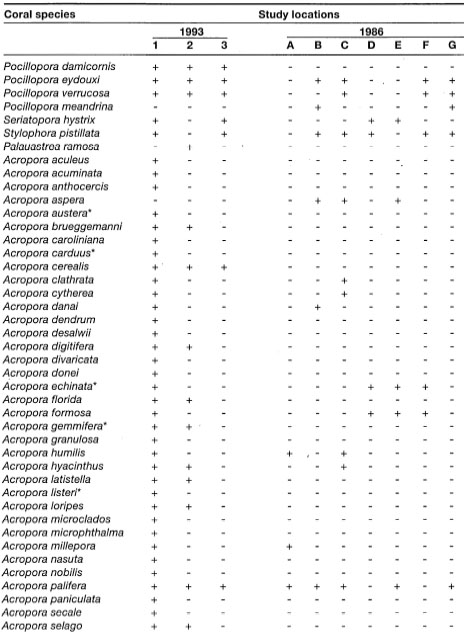

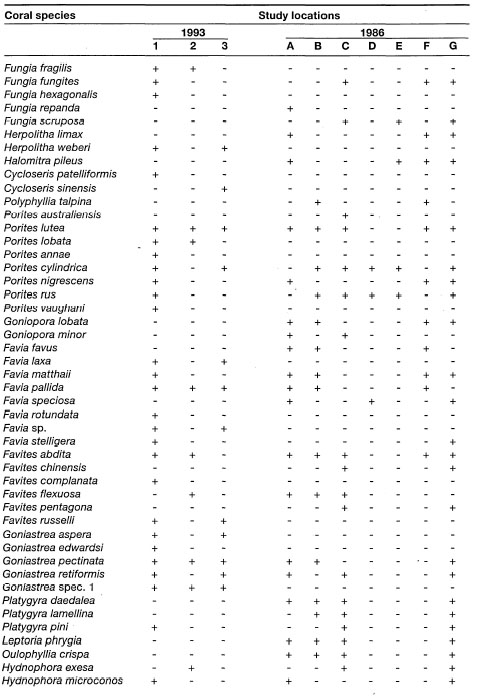
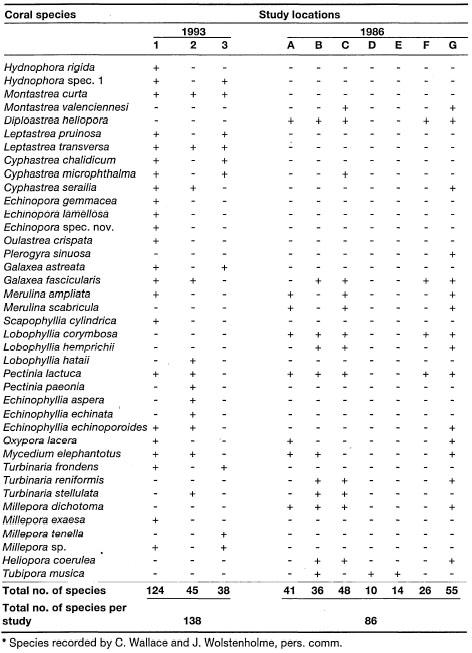

Figure 11.33. Comparison of species richness, Shannon-Wiener's index of diversity (H'c), Simpson's index of concentration (D'c), coral colony abundance and percent live coral cover among the three substrates on Gunung Api in a 1993 study, five years after the eruption. Values represent means per transect, error bars represent standard deviations. Lava: Location 1; Reef: Location 2; Rock: Location 3. Locations as in fig. 11.28.
Source: Tomascik et al. 1996.

Figure 11.34. A five-year-old coral community on the 1988 Gunung Api northern lava flow. Banda Islands, May 1993.
Photo by Tomas and Anmarie Tomascik.

Figure 11.35. Coral community on the original reef substrate not covered by the 1988 lava flow, Gunung Api, Banda Islands. The small size of coral colonies, five years after eruption, suggests that the majority of scleractinians were destroyed during the event.
Photo by Tomas and Anmarie Tomascik.
Figure 11.36. Loose lava rock is the key environmental factor preventing the rapid development of coral community along the southern coast of Gunung Api, Banda Islands. The community is maintained at an early stage of community succession by frequent rock slides associated with unstable substrate and frequent earth tremors. Note the small size of coral colonies.
Photo by Tomas and Anmarie Tomascik.
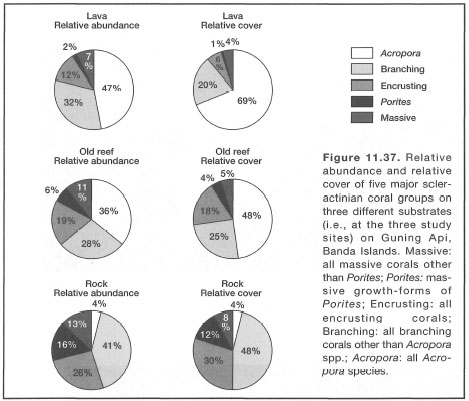
Figure 11.37. Relative abundance and relative cover of five major scleractinian coral groups on three different substrates (i.e., at the three study sites) on Guning Api, Banda Islands. Massive: all massive corals other than Pontes; Pontes: massive growth-forms of Porites; Encrusting: all encrusting corals; Branching: all branching corals other than Acropora spp.; Acropora: all Acropora species.
The acroporids that became established on the lava flow also dominate in terms of coral cover, followed by other branching and encrusting forms. In contrast, small branching, encrusting and Pontes growth-forms were the three most successful colonizers along the southwest coast of the volcano that was smothered by large quantities of tephra, which created an unstable substrate highly prone to slides triggered by frequent earthquakes (fig. 11.38).
One of the most interesting features of the coral communities on all of the lava flows visited during our field studies, was the large size of coral colonies. For example, a few tabulate acroporids on an old lava flow (date not known) on the northwest coast of Siau, Sangihe Archipelago, exceeded 3 m in diameter.(Tomascik and Mah, pers. obs.; Siau 1993). Based on the size frequency distribution of the Gunung Api acroporids, it has been suggested that the radial skeletal extension rates of tabulate acroporids on the 1988 Banda Api lava flow may be as high as 30 cm.yr-1 (Tomascik et al. 1996). These growth rates are among the highest known for this group (fig. 11.39).
Preliminary analysis at McMaster University indicated that linear skeletal extension rates of small Pontes spp. (massive coral growth-forms) on the lava flow can exceed 2.5 cm.yr-1. High nutrient concentrations and high secondary productivity (high zooplankton biomass) of the coastal waters may be one explanation for these rapid skeletal extension rates. During the study, Dr. van Woesik noted that the majority of corals on the lava flow had their tentacles extended during the day. This is a common behavioral response to light for many species of corals, however, some groups are known to have their polyps extended only at night. Within this night coral group, and present on the lava flow, are Caulastrea, Echinopora, Hydnophora, Isophyttia, Montastrea, and Oxypora. On the lava flow, the species with continuously extended tentacles were Acropora cytherea, A. cerealis, A. latistella, A. florida, A. subulata, A. hyacinthus, A. paniculata, A. yongei, Montipora efflorescens, Pocillopora eydouxi, P. verrucosa, and Mycedium elephantotus.

Figure 11.38. South coast of Gunung Api, Banda Islands. A) The rapidly eroding cliffs (i.e., tephra deposits) along some parts of the south coast of the volcano are the 'classic' source of large quantities of sediments that have a significant negative effect on the colonization and survival of coral recruits in the area. B) The loose rock talus along this section of the coastline is the main factor that prevents the development of reef build-up.
Photos by Tomas and Anmarie Tomascik.
It is generally accepted that high concentrations of nutrients, particularly dissolved inorganic phosphorus (PO4-P), inhibit coral calcification rates, and possibly growth rates. Note that reduced coral calcification rates may not necessarily be reflected in reduced skeletal extension rates, but skeletal density will be reduced (Dodge and Brass 1984). At high enough concentrations (>2 μm or μg-at.l-1), phosphorus acts as a crystal poison by inhibiting the formation of CaCO3 crystals (Simkis 1964). The ecological effects of high nutrient concentrations are more difficult to interpret, since other environmental characteristics of the area (e.g., coastal currents, background levels, depth profile, etc.) must be considered. Nonetheless, sufficient data are available to conclude that an increase in nutrient concentrations above ambient will at some point lead to eutrophication. In this discussion we are interested in the effects of nutrient concentrations only slightly above the mean ambient levels, which are usually not investigated in field or laboratory studies. For example, it has been demonstrated that under laboratory conditions, nitrogen and phosphorus have a measurable effect on the shell formation of Tridacna gigas (Belda et al. 1993). Working with juvenile giant clams, they were able to demonstrate that high nutrient levels (N and P) significantly enhanced shell growth rates (i.e., skeletal extension), but at the same time also significantly reduced the weight of the shells. This implies that while the clams are growing rapidly under high nutrient concentrations, their shell densities are much lower. The implication is that lower shell densities may make the tridacnas more susceptible to predation by drilling gastropods, especially during their juvenile stage just after settling from the plankton. However, the nutrient (N and P) concentrations used in the study (i.e., 5-10 μm, N; and 2-10 μm, P) to elicit this response were well above the average ambient levels that the clams are likely to encounter on the reef. Unfortunately, there are no comparable studies using environmentally 'realistic' concentrations. What the study suggests, however, is that similar responses to elevated nutrient concentrations may be elicited in other calcifying photosymbiotic organisms, including the zooxanthellate reef-building corals.
Figure 11.39. Large tabulate acroporids are a dominant component of the 1988 lava flow coral community. This Acropora latistella is being used as a dinner plate by some reef inhabitants, most likely an octopus or a Napoleon wrasse (Cheilinus undulatus). Gunung Api, Banda Islands, May 1993 (maximum diameter 85 cm).
Photo by Tomas and Anmarie Tomascik.
The above-average skeletal extension rates of scleractinian corals, as well as large shell size (maximum size 48 Cm) of Tridacna squammosa (well above the average; R. Braley, pers. comm.) on the Banda Api lava flow, suggests that nutrients may be involved. During the Southeast Monsoon upwelling period, ambient sea surface PO4-P and NOs-N concentrations in the Banda Sea range between 0.11 to 0.56 μm and 0.30 to 3.52 μm, respectively (Institute of Marine Research 1973c; Wetsteyn et al. 1990), and are well above the average values in the Caribbean and elsewhere. During the Southeast Monsoon, all coral reefs in the Banda and Flores Seas are subjected to very similar nutrient conditions (Wetsteyn et al. 1990), with slightly higher levels in the Arafura Sea. This difference is due mainly to the upwelling of 100-150-m-deep Arafura Basin water masses over the Arafura Shelf. According to Wetsteyn et al. (1990), this is an impressive phenomenon that brings nutrient-rich water to benthic and pelagic communities hundreds of kilometres from the shelf break. Nonetheless, coral growth rates comparable to Banda Api corals have not been observed, even on some of the adjacent islands in the Banda Archipelago. This suggests that the effusion of hydrothermal fluids from numerous shallow-water hydrothermal vents around Banda Api may be a significant source of nitrogen and phosphorus to the reef community. For example, it has been demonstrated that rivers draining active volcanic regions in Costa Rica, Central America, carry high loads of dissolved nutrients, which significantly enhance the productivity of the receiving waters (Pringle et al. 1993). Therefore, it is suggested that hydrothermal fluids associated with geothermal processes of the Banda Api volcano may also be a significant source of nutrients. Unfortunately, data to demonstrate this probable connection are not available.
However, thermal springs are a very common phenomenon on Java as well as other volcanic islands throughout the archipelago. Among the best known are the famous thermal springs of Cipanas in West Java, located at the foot of an old lava flow from Gunung Gede (Guntur). According to Hutchison (1981), Gunung Gede's lavas contain between 50%-61% SiO2 and between 0.44%-0.88% KgO, whereas Banda Api's SiO2 content ranges between 62.7%-67.8%, and K2O between 0.59%-0.89%; thus both are classified as andesitic volcanoes. While most lavas from Java volcanoes are quartz normative, the Banda Api lavas are entirely quartz normative (i.e., theoretically highly predictable mineral composition) (Hutchison 1981). It is, therefore, possible that the mineral and nutrient concentrations of the hydrothermal fluids associated with the geothermal activity of these two volcanoes may be similar. Much closer to Banda Api and situated on the Inner Banda Arc are three islands (i.e., Pan tar, Serua and Damar) with active andesitic volcanoes. Hydrothermal activity along the coastline occurs on all three islands, and some of these hot springs were sampled during the Snellius-II Expedition. The expedition visited Banda Api in 1984, but samples from hydrothermal vents were not collected. However, the composition of the Banda Api hydrothermal fluids may be similar to those of Serua and Damar, since the hydrothermal vents are located near the beach (table 11.6). Unfortunately, nutrient analyses for N and P were not done during the Snellius-II Expedition, so possible nutrient budgets cannot be estimated. Therefore, hot spring data from Cipanas are included in table 11.6, since values for P and N can be approximated. The data clearly demonstrate that hydrothermal fluids are significantly enriched in nutrients and other mineral constituents; thus, it is likely that hydrothermal venting will have a measurable impact on the coral communities and the associated reef fauna and flora. Based on our observations thus far, this effect seems to be very beneficial.
The seasonal monsoonal upwelling in the Banda Sea that occurs during the Southeast Monsoon (Wyrtki 1961; Nontji 1975; Birowo and Ilahude 1977; Boely et al. 1990; Ilahude et al. 1990; Wetsteyn et al. 1990; Zijlstra et al. 1990), and the continuous effusion of geothermal fluids from the volcano, are, however, not the only source of nutrients for the Banda Api lava flow coral community. As a result of submarine topography and tidal regime, the Banda Islands also experience daily tidally-induced upwelling events that may significantly influence the nutrient budget over the reef on a daily basis. Based on in situ temperature measurements taken over the reef slope during the upwelling events (with recorded temperature drops from 29.8°C to 26°C), and temperature and nutrient depth profiles taken during the Snellius-II Expedition (Wetsteyn et al. 1990), we have estimated that the upwelling water masses may originate from depths between 50 to 60 m. Nutrient concentrations measured at these depths in the Banda Sea range between 0.130.53 μm and 0.9 - 7.8 μm of PO4 and NOs, respectively (fig 11.40) (Wetsteyn et al. 1990). Note that these nutrient concentrations are not persistent, but occur in pulses.
High ambient nitrate concentrations may, therefore, be one explanation for the recently demonstrated 8 N enrichment of Pontes spp. tissue samples collected directly from the Banda Api lava flow. These values are similar to the δ15N values obtained from corals collected from a sewage-polluted area in the Maldives (Risk et al. 1993). It is worth mentioning that all Acropora colonies were very fragile and brittle, a condition exhibited by Tridacna gigas exposed to high N and N+P concentrations (Beldaetal. 1993).
Coral colonization of lava flows has been previously reported only from Hawaii (Grigg and Maragos 1974), a region with much lower species richness when compared to the Banda Islands, as well as different environmental conditions. The scleractinian fauna of Hawaii is represented by only 12 scleractinian genera and 44 species (Grigg 1983; Dollar and Tribble 1993). In an important ecological study of scleractinian coral colonization rates of Hawaiian lava flows, Grigg and Maragos (1974) clearly demonstrated that Hawaiian scleractinian coral communities smothered by basaltic lava flows may take from 20 to over 50 years (i.e., depending on environmental conditions) to recover to their earlier species richness, percent cover, and diversity levels. The results of the Hawaiian study have been cited, and used frequently, to demonstrate the relatively slow recovery rates of coral communities following major environmental perturbations. These results fit well with the earlier observations made by Umbgrove (1947) at Krakatau. While the coral communities in Hawaii and Krakatau evolved under markedly different climatic and hydrological conditions, they nevertheless colonized a similar (i.e., chemical composition) substrate consisting predominantly of basaltic lava. Different environmental conditions that occur in Sunda Strait (i.e., location of Krakatau) and the Banda Sea (i.e., location of Gunung Api), to a great extent explain current differences in coral community structure and reef development. However, it is suggested that substrate type has been the major environmental factor that has considerably slowed down the reef development at Krakatau. Unlike Gunung Api, where solid andesitic lava is the primary substrate, loose basaltic lava rock and thick tephra deposits that form high eroding cliffs clearly predominate at Krakatau. Thus, coral communities at Krakatau have little chance to become established, a very similar scenario that also exists along the south coast of Gunung Api.
Table 11.6. Major mineral and nutrient constituents of hot springs (hydrothermal vents) from Cipanas (West Java), and Pantar, Nila and Damar on the Inner Banda Arc. The Pantar and Serua hot springs are located on the beach; thus, thermal fluids are contaminated with seawater.

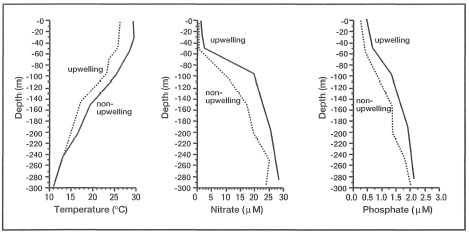
Figure 11.40. Vertical depth profiles for temperature (T°C), nitrate (NO3-N μm) and inorganic phosphate (PO4-P μm), Banda Sea. The stations were in the vicinity of the Banda Islands during the non-upwelling season in April (i.e., Northwest Monsoon), and the upwelling season in September (i.e., Southeast Monsoon). Non-upwelling: April 8, 1970 (Station 16; Cruise No. 1). Upwelling: September 22, 1972 (Station 25; Cruise No. 10).
Source: Institute of Marine Research 1971a, 1973b.
The Gunung Api eruption in 1988 Created a natural experiment where the colonization of an andesitic lava flow could be monitored for the first time. A 1993 study of the lava flow (five years after the eruption) provided first quantitative evidence that, under natural conditions (i.e., unaffected by anthropogenic activities) , coral communities have a remarkable healing capacity to recover from major environmental perturbations (Tomascik et al. 1996). Within five years of the eruption, the new coral community (the embryonic reef) that became established on the lava flow has surpassed the earlier and adjacent coral communities, which were not affected by the eruption, in terms of species richness, abundance, and cover. The rapid colonization and succession may be related to the above-average coral growth rates, and to the apparent preference for andesitic lavas by settling coral planulae. The contrasting results of the Hawaiian and Gunung Api studies are mainly a result of different environmental conditions, frequency of local disturbances (Dollar and Tribble 1993), lava type, biogeographic setting, the diversity of the local and regional species pool, and perhaps anthropogenic influences.
The common feature of most of the young Indonesian volcanoes is that their lavas are mostly andesitic. This seems to be the common denominator in all instances where we have observed vigorous coral reef development to be occurring at the present. Table 11.7 lists the major elements of lavas from all active volcanoes where extensive reef development was recorded. The data presented in table 11.7 clearly demonstrate that major elemental differences exist between the Banda Api and the Hawaiian (Kilauea) lavas.
The 1993 Banda Api study provides sufficient grounds to seriously question the generality of the widely held view that coral reefs do not develop on active volcanoes (Davis 1928). While it is true that Banda Api does not have a continuous fringing reef, long stretches of the coastline, especially along the northeast coastline, are fringed by a well-developed fringing reef. The absence of a well-developed fringing reef along the south and southwest coast of the volcano is primarily a result of unstable substrate, which consists mainly of loose pyroclastics and large lava boulders that are severely scoured by loose volcanic ash particles. However, Banda Api is an integral part of a three-island complex (i.e., Banda Naira, Banda Besar and Gunung Api - not including the smaller islands), which is in fact almost totally surrounded by a coral reef.
Table 11.7. Elemental composition (wt%) of lavas from the Sangihe and the Inner Banda Arc volcanoes that are known to support vigorous reef development.

Figure 11.41. During volcanic eruptions burial of fringing reefs may occur in coastal areas downwind from the eruption. Large sponges such as this Petrosia testudinaria may recover from partial burial. Southwest coast of Ruang Island, Sangihe Archipelago.
Photo by Tomas and Anmarie Tomascik.
High-diversity coral communities, such as those found on the Banda Api lava flow, are not unique to the "Spice Islands". Similar high-diversity and high-cover coral reef communities that now form well-developed fringing reefs have been observed around other active volcanoes in the Banda and Sulawesi Seas (e.g., Ruang Island - with the last eruption in 1949). These observations clearly demonstrate that volcanoes do not have to be extinct or dormant before coral communities, and thus a fringing reef, can develop.
Here again the growth of the reefs was more vigorous than I had expected to find on the slope of an active volcano… Although Gn. Api [north of Wetar] cannot therefore be cited as a proof of the theory that reefs can grow on an active or only recently extinct volcano, it certainly agrees with this hypothesis.—KUENEN 1933
Kuenen's earlier, and often overlooked, suggestion has found strong support in our quantitative study of Banda Api as well as from our qualitative observations on many other volcanic islands with active volcanoes. Thus, contrary to the long-held view, the Gunung Api study in the Banda Islands suggests that intermittent eruptions may actually promote local coral diversity, and that hydrothermal activity associated with geothermal volcanic processes may promote, in an unknown fashion, coral growth rates (Tomascik et al. 1996). Nonetheless, Pelean eruptions without significant lava flows produce considerable amount of pyroclastic "fallout" (i.e., volcanic ash, tuffs, etc.) that may smother considerable sections of coastline, including fringing reef communities (fig. 11.41). Clearly, this area offers exciting new research opportunities that can provide us with much-needed answers to some very complex questions.
Banua Wuhu (Mahengetang) Underwater Volcano
Hydrothermal venting that was observed in Banda Api is not a unique phenomenon in the archipelago, and most likely occurs widely wherever there are active volcanoes. The Galapagos Islands are famous, at least in sport-diving circles, for their gas-bubbling, hot-water vents associated with the volcanic nature of the islands. The same phenomenon, but on a much grander scale, was observed on the Sangihe Ridge, which stretches from North Sulawesi to Mindanao in the Philippines. The ridge is a volcanic arc delineating the western border of the Molucca Sea, and is part of a unique arc-to-arc collision complex (Jezek et al. 1981). The volcanic arc, about 550 km long and up to 70 km wide, has a series of volcanoes, spaced 30-90 km apart, that lie between 130-180 km above the centre of a west-dipping Benioff zone that is about 650-km-deep (Jezek et al. 1981). More than 20 of the islands, along the southern and central section of the volcanic arc, are young Quaternary volcanoes. All of these have well-developed fringing reefs that in most instances encircle the entire island, with flourishing coral communities characterized by high diversity and cover. In all, there are 13 active volcanoes along the ridge, including an active submarine volcano. The southernmost volcano on the Sangihe volcanic arc is Unauna, in the Gulf of Tomini, which last erupted in 1983. Most of the islands in the northern region of the arc (i.e., north of Sangihe Island) are capped by thick coral reef deposits, in addition to being currently surrounded by extensive fringing reefs (e.g., Kawio Islands).
Box 11.4. Corals from the Banda Sea as environmental recorders.
J. Heikoop and M. Risk, Department of Geology, McMaster University, Hamilton, Ontario, Canada.
During recent years, the flourishing reefs of the Banda Islands, Indonesia, have been affected by the 1988 eruption of Banda Api and have experienced oceanographic variability associated with their presence within the Western Pacific Warm Pool. Massive corals collected from reefs offshore of the Banda Islands contain physical and chemical records of these phenomena in their aragonite skeletons.
Banda Api is a small volcanic island forming part of the Banda Arc, a collision zone between the Eurasian and Indo-Australian tectonic plates. The volcano last erupted May 9, 1988, depositing up to 1 m of ash on the surrounding coastline. Local coral mortality resulting from the eruption was extensive, particularly among branching species. In May, 1992, four heads of the coral Porites lobata were collected just offshore of Banda Api to determine if a record of this eruption might be contained within their skeletons. It was thought that ash from the eruption may have been incorporated into the skeletons and that this material would be revealed by petrographic investigation.
When the coral heads were cleaved open, two interesting features were immediately evident. All four corals contained extensively bioeroded death/regrowth surfaces through which volcanic ash had infiltrated into the skeletons (fig. 11.42). At the same level within the skeleton as the death surfaces, a thin, black, iron-rich precipitate we have termed the "Banda Band" was also found (fig. 11.42). This band quickly oxidized to an orange colour upon exposure to the atmosphere. Using X-radiography and stable isotope chronologies, the contemporaneous Banda Band and death surfaces in each coral were found to have formed at the time of the May 1988 eruption of Banda Api (Heikoop et al., in press).
Death surfaces formed over parts of the skeleton where ash from the eruption could not be quickly cleared away by wave energy or by coral sediment rejection. These surfaces were subsequently bioeroded and infiltrated by small ash particles prior to tissue expansion and regrowth over damaged areas of the coral colony. The origin of the Banda Band is thought to be related to local hydrothermal activity which has been ongoing near the collection site since the eruption. A large area of hydrothermal venting deposits iron oxides on surrounding rocks and sediments close to the collection site. It is likely that enhanced activity, coinciding with the 1988 eruption, was responsible for deposition of the chemical precipitate onto the coral skeletons.
It is unlikely that any discrete record of the 1988 eruption will be preserved within surrounding sediments, due to the relatively moderate size of the event and the occurrence of extensive biological and hydraulic sediment reworking. It is quite possible, however, that century-old massive heads in this or other similarly active volcanic/hydrothermal areas may record one or more of these geologically instantaneous events and that similar records could be found in fossil corals.
The hydrothermal venting around Banda Api has also been of interest as a source of possible nutrient input to the reef system. This work has focused on a number of coral tissue samples collected from small colonies of Porites lobata growing directly on the Banda Api lava flow. These corals had quickly colonized the lava flow which had entered the sea on the north coast of the island during the 1988 eruption (see Banda case study this section). Hydrothermal venting occurs adjacent to this lava flow. The coral tissues were analyzed at McMaster University and were found to be enriched in the heavy isotope of nitrogen. They were, in fact, among the heaviest tissues (δ15N=6.8 ±0.5 %0; n=9) from among the more than 200 samples we have measured from reefs around the world. Carbon isotopic values were typical of other shallow-water corals we have measured.

Figure 11.42. Photo of a cleaved coral head collected offshore of Banda Api. Large arrow indicates "Banda Band". Small arrow points to bioeroded death surface through which ash (a) has infiltrated into the skeleton. Scale 1:2.

Figure 11.43. δ18O record reconstructed from a coral colony collected off the west coast of Run Island in the Banda Sea. Small drilled samples were taken sequentially down the major growth axis of the coral and analyzed on a VG SIRA mass spectrometer to obtain this record. Timing of the record is indicated at the top of the graph. Chronology was established by X-radiography.
Heavy nitrogen isotopic signatures in corals may reflect high light levels (Muscatine and Kaplan 1994; Heikoop et al., in prep.), high nutrient concentrations or an isotopically enriched source of dissolved inorganic nitrogen (DIN) (Risk et al. 1993). All of these factors may be important in this particular setting. Higher concentrations of DIN or an isotopically enriched nitrogen source could be provided by local hydrothermal venting. It is also possible that these corals may be affected by the endo-upwelling mechanism proposed by Rougerie and Wauthy (1993). An active volcanic system such as Banda Api could be expected to create a geothermal gradient, causing upward convection of deep oceanic waters through the volcanic substrate upon which the reef is constructed.
Reefs developed around the Banda Islands are not only influenced by volcanism. These reefs are also flourishing in the warmest body of water on Earth, the Western Pacific Warm Pool, with temperatures generally greater than 28°C (Yan et al. 1992). During El Niño/Southern Oscillation (ENSO) events the warm pool migrates eastward to be replaced by somewhat cooler waters. ENSO events are also associated with severe drought in Indonesia. Given the severe impact of ENSO on eastern Pacific sea surface temperatures and meteorological conditions around the globe, proxy records of this pan-Pacific phenomenon are needed for climate reconstruction modeling. Records from the warm pool are of particular importance in this regard, as sea surface temperature and salinity anomalies in this body of water are potentially responsible for causing the westerly wind bursts which signal the onset of an ENSO event (Enfield 1989).
An oxygen isotope record from a Pontes lobata skeleton is presented in figure 11.43. This preliminary work, from a young coral colony collected at Run Island, shows several features which may be of importance. The trend towards slightly lower δ18O between 1984 and 1990 could be due at least in part to a warming trend in the warm pool (Heikoop and Risk 1994). Evidence for warming is contained in both satellite data (Yan et al. 1992) and ship-collected COADS data for this time period. The isotopic trend is interrupted in 1991-1992, which was the beginning of a large prolonged ENSO event. The slightly higher δ18O values associated with this event could be due to a combination of lower SST and increased δ18O of seawater associated with drought conditions (Heikoop and Risk 1994). However, Yan et al. (1992) point out that significant cooling may have occurred as a result of the 1991 eruption of Pinatubo in the Philippines, which released large quantities of aerosols into the stratosphere, thus reducing insolation. There is only very weak evidence for the presence of the moderate 1986-1987 ENSO event in this record. The isotopic record does not extend back far enough in time to record the severe 1982/1983 ENSO event. Isotopic reconstructions from longer coral records have the potential to greatly increase understanding of ENSO in this area.
Corals are proving to be very important proxy recorders of both volcanism and oceanographic variability in the marine realm of the Banda Islands. With proper calibration, such records could be of great help in filling in gaps in our knowledge of past natural phenomena associated with this region.
The Sangihe Ridge has a high diversity of coastal and oceanic habitats, such as, for example, Pasige Island, which is an oceanic platform reef (almost an atoll - but not quite) with an extensive mangrove forest in one part of the shallow (<5 m) lagoon - a unique environment, indeed. The habitat diversity of the volcanic arc easily surpasses that of the Great Barrier Reef, and supports an impressive diversity of reef-associated flora and fauna - not yet studied. A qualitative account (i.e., based on field and video records) of scleractinian coral species richness in the area ranges from 180-250 species, with higher species richness in the northern sections of the ridge. The chain of volcanic islands along the Sangihe Ridge may serve as an important bridge, or a series of stepping stones (Veron 1995), between the Indonesian and the Philippine centres of coral diversity. This may also be an area where significant overlap of coral reef fauna and flora between the two centres may be expected to occur. The predominant current direction is southwards, thus the Sangihe Archipelago can be considered as down-current from the Philippines almost year-round (Wyrtki 1961). A particularly strong connection may exist between the faunas of Mindanao to the north and Sangihe and Kawio Islands in the northern regions of the volcanic arc. Unfortunately, this diverse region of the Indonesian Archipelago has been generally ignored by the scientific community, even though the latitudinal distribution patterns of coral reef fauna and flora may have important biogeographical implications.
One of the most fascinating environments on the volcanic arc is Banua Wuhu, the only active submarine volcano in Indonesia, which fortunately is easily accessible by scuba. The slightly convex summit of Banua Wuhu is about 500 m in length, along a southwest-to-northeast axis, and is about 250 m wide with an average depth between 6 to 10 m below sea level. The summit of the volcanic peak (about 13 ha) is covered by large volcanic rocks heavily encrusted by calcareous algae, bryozoans and sponges. In many areas where the rocks are coated by a layer of iron-oxide precipitate (fig. 11.44), macroalgae predominate.
The summit is a barren zone as far as the coral fauna is concerned, since very few rocks have been colonized by scleractinians. Here and there are a few, usually bleached, colonies of Pocillopora damicornis and Stylophora pistillata; however, encrusting Millepora spp. seem to be abundant, as was painfully apparent on a number of occasions.

Figure 11.44. Banua Wuhu submarine volcano located just off Mahengetang Island, Sangihe Archipelago, North Sulawesi. The summit of the submarine volcano is characterized by geothermal activity and continuous gas venting.
Photo by Tomas and Anmarie Tomascik.
Extensive gas venting is occurring throughout the summit area, with streaks of bubbles rising to.the surface. At the periphery of the volcanic summit, coral abundance increases as a number of coral species have colonized suitable substrates, but the abundance is very low and percentage cover is well below 1%. The dominant coral species that were able to colonize this barren and inhospitable environment are mainly small (rarely more than 10 cm in maximum diameter) encrusting forms of Goniastrea pectinata, G. australensis, Montipora efflorescens, Leptoseris spp., Favites spp., and small, stubby, wave-resistant growth-forms of Pocillopora damicornis, Stylophora pistillata, Montipora digitata, Acropora gemmifera, A. humilis and A. digitifera. In comparison, Millepora spp. form larger colonies that in some places cover a considerable area. This assemblage of coral species is not surprising, since the summit of the volcano is swept by tidal currents with velocities of up to 4 m.sec-1.
The western slope of the volcano consists mostly of seemingly solid lava substrate. However, there are sections where loose volcanic rock predominates, and this area is generally dominated by encrusting calcareous algae (mainly Porolithon spp.), corals (Leptastrea spp., Cyphastrea spp., Favites spp., Pseudosiderastrea, Gardinoseris spp., Oxypora spp.), Millepora spp., sponges and bryozoans. However, the solid substrate was covered with a luxuriant, high-diversity coral community (>100 species) to a depth of about 30 m (fig. 11.45), where a sharp transition from solid substrate to loose volcanic rock occurred. The most noticeable feature of the west slope assemblage was the relatively small size of coral colonies, indicating that this may be a relatively young community, most likely less than 10 years old.

Figure 11.45. The west slope of Banua Wuhu submarine volcano. Diverse coral assem-blages occur in areas where viscous andesitic lava provides a solid substrate, not prone to rock slides during frequent tremors.
Photo by Tomas and Anmarie Tomascik.
Local reports indicate that there may have been some seismic activity in the area in 1991, which may explain the higher abundance of smaller-diameter tabulate acroporids. However, some large Pontes spp. colonies were present. It seems that frequent shocks and tremors are a common occurrence, and a number of large Pontes spp. colonies located about 650 m to the northeast of the summit showed clear fracture lines through the colony, which were, however, overgrown by new tissue (fig. 11.46). During our brief visit, loud, thunder-like, rumbling sounds were heard during the dives, indicating continual volcanic activity.
In contrast to the west slope, the east slope of the volcano consisted mainly of unconsolidated volcanic rocks. Most rocks were covered by a thin layer of red precipitate, which in some areas inhibited the colonization process by encrusting organisms, which were not abundant. However, the bottom surfaces as well as sides of most overturned rocks were heavily encrusted with calcareous algae and other encrusting organisms. This suggests that substrate mobility is high, which in turn may explain the low abundance of corals. The entire east slope was almost devoid of any large-size coral colonies; nonetheless, most of the volcanic rocks were encrusted by coralline algae (e.g., Porolithon spp., Lithothamnion spp. and Neogoniolithon brassica).
The algal-dominated substrate on the east slope may account for the presence of large-feeding-aggregations of herbivorous reef fish which graze in the area (e.g., Acanthurus fowleri, A. nigricans, A. nigricauda, A. japonicus, A. leucocheilus, Scams frenatus, S. globiceps and S. psittacus) (fig. 11.47). In contrast, the west slope had a significantly higher abundance of planktivorous reef fish (e.g., Pseudanthias dispar, P. pascalus,

Figure 11.46. In areas with intense seismic activity, massive coral colonies, such as this Pontes lobata (located on the west coast of Mahengetang Island about 650 m northeast of the Banua Wuhu summit), are susceptible to earthquakes and earth tremors. Earthquakes in this area occur frequently, and cracks in coralla of massive species are not uncommon.
Photo by Tomas and Anmarie Tomascik.
Luzonichthys earlei, Acanthurus thompsoni, Paracanthurus hepatus, Heniochus diphreutes, Hemitaurichthys polylepis) as well as a higher abundance of reef piscivores (i.e, Epinephelus spp., Cephalopholis spp., Gracilla albomarginata, Plectropomus spp., Lutjanus spp.) and reef-associated pelagic species (e.g., Caranx sexfasciatus). The higher abundance of planktivorous reef fish on the west slope may be related to higher abundance of demersal zooplankton, which may be a result of greater substrate heterogeneity offered by luxuriant coral growth (fig. 11.47). Greater abundance of piscivores is also related to greater habitat complexity associated with coral growth.
Shallow-Water Hydrothermal Vents
A major shallow-water hydrothermal vent area is located on the northwest section of the volcano at a depth of about 10 m. Within a 100 m periphery of this area, the loose volcanic rock substrate is covered by red iron-oxide precipitate with little or no biota visible. While the entire area is actively venting, vigorous and continuous release of hydrothermal fluids and gas also occurred from a large solid ejectamenta about 5 m in diameter and 10 m high. The solid volcanic rock had numerous fissures (up to 5-10 cm wide) from which large volumes of hydrothermal fluids (with temperatures between 51°and 53°G) and gas were being continuously and forcefully ejected (fig. 11.48).
The salinity of the hydrothermal fluid was 34%o-35%o, indicating strong contamination by seawater. The surrounding substrate was covered by a layer (1-2 mm thick) of dust-like, red iron-oxide precipitate, which is surprising, considering the high current velocities and strong wave action in the area. The substrate immediately to the northeast of the vent consists of unconsolidated oolitic iron nodules, which are concentrated in small 'pools'. The topography of the roughly 50 m2 area had a distinct desert-like appearance.
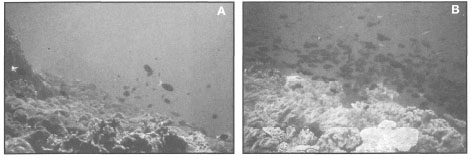
Figure 11.47. Visual comparison of the habitat type and reef fish assemblages between the east (A) and west (B) slopes of Banua Wuhu underwater volcano, Sangihe Archipelago.
Photos by Tomas and Anmarie Tomascik.
Just outside this relatively barren area is a 20-50-m-wide zone where the substrate is heavily overgrown by dense mats of macroalgae, which grow in such profusion that they outcompete all corals and calcareous algae. The dominant macrophytes were Codium edule, Zellera tawallina, Lyngbya majuscula, Galaxaura cf. lapidescens, Tricleocarpa cylindrica, Amansia fraxinifolia, and Spyridia filamentosa. This algal community is consistent with high nutrient concentrations in hydrothermal fluids.
Interstitial water temperatures taken at a depth of 10 cm below the sediment/water interface were 39°-42°C, while at the sediment/water interface temperatures were 30.5°C, and at 0.1 m above the sediments temperatures were 28.8°29.2°C, which were the ambient levels. It seems that in addition to the probable high nutrient concentrations associated with the hydrothermal fluids, above-ambient seawater temperatures beneath and at the sediment/water interface may be a significant factor contributing to algal monopolization of the substrate. Strong currents and heavy wave action are rapidly mixing the hydrothermal fluids with seawater, creating a localized high-nutrient environment.
During our survey of this remarkable place, an exciting new discovery was made in the shallow coastal waters of Mahengetang Island, located about 400 m to the east of Banua Wuhu. Mahengetang Island and Banua Wuhu (fig. 11.49) comprise a single volcanic complex located at the western edge of the Sangihe Ridge. While the western margin of this volcanic complex rises steeply from a 5-km-wide and 500-m-deep channel that separates Mahengetang from Kahakitang Island to the northeast, the north, east and south margins of the volcanic complex plunge to depths greater than 1000 m only a few hundred metres offshore.
Banua Wuhu and Mahengetang are separated by a narrow (350 m) channel with a maximum depth of about 80 m. Mahengetang Island is surrounded by a continuous fringing reef approximately 4.8 km in length. The lack of fresh-water streams or rivulets on this roughly 6 km2 heavily vegetated volcanic island may account for the uninterrupted nature of the reef. We were informed by the transient fishermen on the island that fresh-water supplies must be brought in from the adjacent islands, since the ground water on Mahengetang apparently lacks a shallow fresh-water lens. The fringing reef that surrounds the entire island is on average about 200 m wide, but at the southwest coast the reefal area extends some 400 m offshore. During our survey of the reef along the northwest coast of the island (about 100 m offshore), we came upon a section of the reef that was dominated by massive colonies (many >1.5 m in diameter) of Pontes lobata, Pontes lutea, Pavona minuta, Pavona clavus, Heliopora coerulea and Millepora spp., but only a few branching species were present (i.e., Pontes cylindrica) (fig. 11.50).

Figure 11.48. Banua Wuhu submarine volcano located just off Mahengetang Island, Sangihe Archipelago, North Sulawesi. The volcano is actively venting gas through numerous fissures at the summit, which is located at a depth of about 10 m. The composition of gas is not known, however temperatures of the hydrothermal fluids exceed 53°C.
Photo by Tomas and Anmarie Tomascik.

Figure 11.49. Location and bathymetry of Banua Wuhu and Mahengetang Island, Sangihe Archipelago. Soundings in metres. White star location of ooid pools. Black star location of major hydrothermal venting on Banua Wuhu.
From Hydrographic Chart No. 183.
Unlike other areas of the fringing reef, where carbonate fragments were the main sediment components derived from the diverse reef community (e.g., corals, algae, molluscs, echinoids, etc.), the sediments in the massive coral habitat were made entirely of loose lava rock, and extensive "pools" of unconsolidated dark red-brown oolitic iron nodules (ironstones), that earlier were observed on Banua Wuhu (fig. 11.51).
Next to Banua Wuhu, the unconsolidated oolitic "pools" at Mahengetang were the main areas of hydro thermal activity, and totally devoid of any carbonate material. This is difficult to explain, since these "pools" are in the midst of a flourishing fringing reef. The coral community at Mahengetang Island occupies a coastal area roughly between 1.2 to 2.0 km2, extending from the shore to the 30 isobath. In the northwest sector of the reef there was, however, almost total absence of branching coral species, thus the dominance of massive forms which do not break or fragment easily may partially explain this phenomenon, but not entirely. The reef to the south, with up to 100% live coral cover, is dominated by branching (e.g., Seriatopora hystrix, S. caliendrum, Acropora yongei, A. formosa, A. nobilis, A. aspera, Pontes cylindrica, P. nigrescens), laminar (Acropora hyacinthus, A. paniculata, A. cytherea), and foliaceous (Montipora aequituberculata, Echinopora lamellosa, Pachyseris spp. Merulina ampliata, Pectinia spp. Turbinaria spp., and Pavona cactus) forms. The sediments were a mixture of reefal carbonates and volcanic sands, but with carbonates dominating. The substrate itself was carbonate, but we do not know how thick. This contrasts with the oolith area where the substrate consists of loose volcanic rocks overgrown by massive corals (e.g., Pontes spp.) and Millepora spp.
Preliminary geochemical analyses, being conducted by Jeff Heikoop at McMaster University, indicate that these unconsolidated sediments are a type of ferriferous ooliths (ooids/pisoids), made entirely of concentric rings of iron oxide that have grown around a volcanic fragment or some other volcanic particle (fig. 11.52).
The ferriferous nodules are roughly spheroid to ovoid in shape, with diameters ranging between 1-4 mm. The oolites are made of concentric accretionary layers of precipitated iron, each layer averaging about 0.05 mm in thickness. The inner core of the nodules consists of goethite, while the outer layers are made of amorphous iron (Heikoop et al., in press). The mineral composition of the ooliths is 75% iron oxide including the rock core. This high percentage of iron is used by mineralogists to identify iron ore deposits. Thus, we may have stumbled onto an iron ore deposit in the making. There is, however, no evidence of any carbonate content whatsoever. Similar types of sediments form economically-important Jurassic iron ore deposits in England and other parts of Europe. However, their origin is still an enigma. The structure of the iron ooliths from Mahengetang is new for contemporary minerals, and work is being continued to solve this new exciting scientific puzzle. It is widely believed that iron ooliths originate through replacement of calcareous ones by iron oxide (i.e., they lack primary origin). The findings from Mahengetang are a serious challenge to this hypothesis, since the formation of iron-oxide-rich (iron ore grade) ooliths clearly demonstrates that the ancient examples most likely had a primary origin.
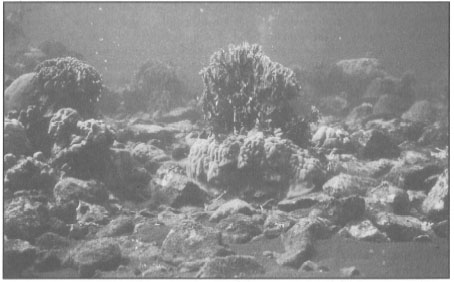
Figure 11.50. Massive Porites spp., Heliopora coerulea and Millepora spp. are the dominant coral growth-forms along the margins of actively venting hydrothermal gas fields on the west coast of Mahengetang Island. Note gas bubbles rising to the surface at front left.
Photo by Tomas and Anmarie Tomascik.

Figure 11.51. Large colony of unattached Porites lobata growing in a field of ferriferous ooids.
Photo by Tomas and Anmarie Tomascik.
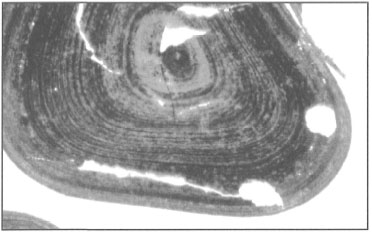
Figure 11.52. A thin section of Mahengetang ferriferous ooid/pisoid showing concentric rings of iron oxide.
Courtesy of J. Heikoop, McMaster University.
Surprisingly, the iron ooliths were not found in Banda Api, or around any of the other volcanoes that were surveyed. It seems, however, that these peculiar structures, tentatively named ferriferous ooids by Jeff Heikoop, may be products of submarine volcanism, most likely associated with hydrothermal and gas vent areas in deeper water (>5 m). The hot vents in Banda Api became active only since the 1988 eruption, and therefore, they are relatively young, even though they most likely contain a significant amount of iron in solution. Furthermore, the main hydrothermal vent area in Banda Api is very shallow (from the beach to a depth of <1 m) and located along the exposed southwest coast of the volcano. Thus, much of the material being precipitated in this area gets washed upon the shore. Since the shoreline deposits are mainly loose tephra, rapid erosion is occurring and new material is being continuously brought into the area. Nonetheless, most of the intertidal and subtidal rocks in the area are covered by a layer of iron-oxide precipitate, which is also accumulating in the sediments. It seems, however, that the overall abiotic environmental conditions on the southwest coast of Banda Api may not be conducive to oolith formation, even though the primary ingredients seem to be there. In addition, a biogenic role in the formation of the ferriferous nodules cannot be ruled out. Using a SEM (scanning electron microscope), Jeff Heikoop found that the oolith samples from Mahengetang are covered by a layer of benthic diatoms (single species not yet identified). It appears that the diatoms are attached to the ooliths by mucus in which many of them become encapsulated. If benthic diatoms play a major role in the formation of these structures, then light and nutrient conditions become very important.
In comparison to Banda Api, the large hydrothermal vent area in Mahengetang where the ooliths form wide "desert-like plains" or "pools" is located on a lee side of the island at a depth of about 7 m, thus, relatively protected from the weather. On the wave-exposed Banua Wuhu, the oolith deposits occur in deeper water (<15 m) and on the lee side of the large ejectamenta where they are protected from considerable wave action that takes place in the shallow water. However, the tidal currents in these two areas can reach 2.5 m.sec-1 or more. With regards to Banua Wuhu, the oolith sediments may not be indicative of place of origin, but rather they may constitute a deposit. The environmental conditions seem to fit the theoretical requirements for oolith formation, since oolitic limestones apparently display current-bedding. Iron precipitation is most likely a result of mixing of iron-rich hydrothermal fluids with seawater at the sediment/water interface. It is probable that the continuous gas venting through the sediments and the relatively steady and fast currents are providing the necessary agitation which promotes the formation of the 'oolitic structures' (fig. 11.53).
This may well be the case, since it was observed that when the gas is being released through the sediments, the top layer seems to float just a fraction above the rest of the sediments. The actual venting (i.e., release of gas) lasts only about 5-10 seconds, followed by a 5-minute build-up phase, before the next venting. Unfortunately, the chemical composition of the gas is not known.
The unconsolidated oolith layer seems to act as a semipermeable barrier for the gas whose pressure has to build up to a certain level before being forcefully released through the sediments. The gas venting is synchronized and highly periodic with an on-off cycle of about five minutes, at the time of our visit. As a result of the synchronization, the gas is released at the same time from the entire area, which is up to 100-200 m2, and one has the impression of being in a giant aquarium with thousands of air stones producing air bubbles. As a simple analogy, the effect of the gas venting is similar to back-flushing a sand filter. The oolitic sediments are up to 0.5 m thick (likely deeper), and the subsurface water about 20 cm below sediment/water interface has a temperature of 37° to 42°C, indicating strong geothermal activity below the fringing reef. The temperature differences are most likely related to tidal pumping, since higher temperatures were measured at ebb flow. A similar phenomenon was observed at Banda Api, where a large intertidal hydrothermal vent at the base of the 1988 lava flow was venting only during ebb flow, with temperatures reaching 47°C in the shallow intertidal pool adjacent to the vent.
It seems quite amazing that the hot water just a few centimetres below the sediment/water interface, as well as the continuous gas venting, does not seem to have a negative effect on the coral community. Numerous Pontes lobata and P. lutea colonies were observed in the middle of some of these oolitic fields without any signs of stress; quite the opposite, they looked very healthy even though they were in the midst of a seemingly alien environment.

Figure 11.53. A) Section of a large (30-50 m2; >50 cm deep) deposit of ferriferous ooids/pisoids on the west coast of Mahengetang Island. Note the absence of calcareous material and the homogeneous nature of the ferriferous deposit. B) The extensive ooid/pisoid fields are major hydrothermal and gas venting areas.
Photos by Tomas and Anmarie Tomascik.
EARTHQUAKES
Earthquakes are a common occurrence in many parts of the world. Examination of the main active earthquake belts reveals that, almost without exception, major earthquake activity occurs along the margins of interacting tectonic plates, while they rarely, if ever, occur in many other regions. On a geological time scale the Indonesian Archipelago is relatively young, formed by the interaction of three major tectonic plates, the Australian-Indian Plate to the south, the Pacific Plate to the east and the Eurasian Plate to the north, as well as a number of smaller plates (see chapter 2). Major interactions occur along all active margins, for example, along the Sunda Arc where the Indian-Australian Plate is being subducted under the Sunda Shield portion of the Eurasian Plate. Thus, Indonesia is one of the most earthquake-prone regions in the world. Fortunately, the most destructive earthquakes (i.e., shallow earthquakes) have their epicenters located in offshore oceanic regions, or in relatively underpopulated areas (Katili 1985), otherwise the damage sustained would be much greater. However, with the predominant offshore location of major earthquake epicenters, the probability of a tsunami impacting upon the shorelines increases as well. According to Katili (1985), 10% of the world's earthquake activity occurs in the Indonesian Archipelago, whose total area of 5.8 x 106 km2 (including the Exclusive Economic Zone - EEZ) (Uktolseya 1990) represents roughly 1.1 % of the earth's surface. Table 11.8 provides a summary of the Indonesian seismo-tectonic zones.
As was clearly pointed out in the RePPProT (1990) report, nearly 90% of the Indonesian population lives in regions where earthquakes with a magnitude of at least 8.0 on the Richter scale can be expected, and Jakarta is within one of these danger zones. The disastrous consequences of the earthquakes in Managua, Nicaragua, and Mexico City, Mexico, two cities built in areas with not-too-dissimilar geologic histories, should not be forgotten. Recent tragedies in Flores and South Sumatra serve to remind us of the dynamic nature of the archipelago.
Shallow-water earthquakes (earthquake foci <60 km deep) occur primarily along the Great Sunda and Banda Arc subduction zones as well as along known transcurrent faults (e.g., Great Sumatran fault zone, as well as the Gorontalo, Sorong or Palu-Koro-Matano fault zones) (Katili 1985). Earthquakes with foci at a depth between 100-300 km occur along the entire length of the archipelago on either side of the Great Sunda and Lesser Sunda volcanic arcs.
Deep earthquakes with foci between 500-800 km occur primarily along an east-west axis between the Java and Banda Seas, as well as along a north-south axis between Sulawesi and Mindanao, Philippines (Katili 1985). Based on the location of the deep earthquake epicenters, Katili (1985) concluded that the Wadati-Benioff zone north of Java has penetrated the lithosphere to a depth of about 700 km.
Table 11.8. Indonesian seismo-tectonic zones and magnitude of seismic activity.
Coral Reefs and Earthquakes
A brief look at figure 2.11 reveals that some of the most diverse coral reef communities have developed in one of the world's most earthquake-prone regions. We are all well aware of the destructive nature of earthquakes on land, however, very little information is available on their effects in shallow-water coastal areas. Comparing figures 11.6 and 2.11 begs the question whether frequent earthquakes in Indonesia have a significant influence on coral community structure as do the cyclones on the Great Barrier Reef. Obviously, the impacts of earthquakes and cyclones on coral reefs differ greatly, and thus different responses should be evident. Unfortunately, no quantitative information exists on this topic. However, some qualitative observations suggest that, at least on a local scale, earthquake activity may have an important influence on community structure and reef zonation patterns.
Coral Reefs in Sunda Strait
Coral communities around Sertung, Rakata and Rakata Kecil Islands (i.e., Krakatau Islands) have colonized relatively unstable substrates, ranging from coarse loose sand (e.g., Sertung Island) to loose, but large, basaltic rocks (Rakata and Rakata Kecil). Reef slopes facing Anak Krakatau are usually gentle and sheltered. The seaward-facing slopes are steeper and exposed to the seas. It seems that the slow recovery (i.e., on a local scale) of the Krakatau coral reef fauna is directly related to the general absence of suitable substrate, and the unstable nature (i.e., loose rock and earthquakes) of the existing basement. Frequent earth tremors in the area are a significant factor influencing coral community structure in the area. Particularly sensitive to earth tremors are the tabulate acroporids such as Acropora hyacinthus, A. cytherea, and A. clathrata as well as some large corymbose species such as Acropora latistella and A. valenciennesi. Overturned colonies belonging to these species are a common sight in the area. High abundance of small tabulate acroporids suggests that earth tremors may be frequent, and of sufficient magnitude to cause breakage of colonies that are > 50 cm in diameter.
The significant role of earthquakes, or earth tremors, in structuring coral communities in Sunda Strait was demonstrated to us during one of our field surveys in the area. Studies focusing on the effects of earth tremors have so far not been conducted in Indonesia, even though in tectonically-active regions such as the Sunda Strait they may be a significant factor. Peucang Island is located at the southeast entrance to Sunda Strait, along the protected northwest coast of Ujung Kulon Peninsula, about 70 km south of the Krakatau Islands. The entire coastline of Peucang Island (12 km) and Ujung Kulon, from Tg. Layar to Tg. Alangalang (26 km), is fringed by a healthy fringing reef with a reef flat that extends between 50 to 100 m offshore. In most places, coral communities on the reef slope extend to a maximum depth of about 10 m, thus the 20 m isobath, about 800 m offshore, can be used as the maximum outer limit of reefal habitats which cover about 30 km of coastline. This is a unique region, since along the 26 km of coastline the fringing reef seems to merge with a primary tropical rain forest that covers the entire Ujung Kulon Peninsula. There are not many places where the fauna and flora of the two most diverse ecosystems on the planet (i.e., rain forest and coral reef) can be studied side-by-side; thus, while studying the behaviour of Amphiprionocellaris on the shallow reef slope (false clown anemonefish), one may also observe the behaviour of Buceros rhinoceros (rhinoceros hornbill) high in the overhead canopy of the rain forest (Tomascik and Mah, pers. obs.).
Sukarno and Suharsono (1983) recorded 30 coral genera represented by 65 species, with Acropora brueggemanni being the dominant, during their survey of Peucang Island. However, the exact location of the survey was not given. During our 1992 survey of two sites on the southeast coast of the island, facing Ujung Kulon, the most abundant species was Pontes cylindrica that formed large monospecific stands, and Acropora brueggemanni was not recorded. This may be due to significant local differences in community structure among the various habitats around Peucang Island. However, among the common acroporids in the 1992 survey were Acropora hyacinthus, A. formosa, A. nasuta, A. nobilis, A. microclados, A. pulchra, A. latistella, A. yongei, and A. yongei. A unique feature of this acroporid assemblage was that all colonies were lightly calcified (R. van Woesik, pers. comm.), and even in sheltered areas the colonies were not large. Only a few Acropora hyacinthus plates were greater than 50 cm in diameter, even though colonies of this species are known to attain 2-3 m diameters. Most other laminar species were also <50 cm in diameter.
During a 1991 visit to the area, the first impression upon arriving at the reef site was that a bleaching event, or an Acanthaster planci outbreak, had hit the area. Upon closer examination, however, we found that the numerous white blotches on the shallow reef flat (<3 m) and the reef slope were mostly overturned or toppled colonies of A. hyacinthus, plus a few Acropora latistella. Turning over numerous toppled colonies, it became apparent that all were still quite healthy, indicating that whatever caused this massive physical impact (over 30 colonies were counted) occurred relatively recently. All toppled colonies had diameters greater than 50 cm. Examining the base by which the corals were attached to the substrate, revealed a sharp clean cut. This type of damage is believed to be associated with earth tremors in the area. Because the coral skeletons are less calcified, most likely a function of the low-energy environment in this area, the base of the corallum is weaker, thus more easily susceptible to substrate mobility associated with earth tremors. The absence of large-diameter (i.e., >1 m) Acropora hyacinthus seems to support this contention. The size distribution of A. hyacinthus in this area may therefore be controlled by earth tremors. This may have a significant effect on the overall community structure, since Acropora hyacinthus is an effective competitor, with a significant shading effect. Thus, periodic earth tremors of sufficient magnitude in this area seem to be exerting significant selective pressure on a specific growth-form. The effects on the community structure are most likely very different from those of tropical cyclones in other coral reef regions.
Earthquakes and Banda Api Eruption
Earthquakes, however, are not unique to the Sunda Strait. During volcanic activity, frequent and intense earth tremors usually precede and sometimes follow the eruption (fig. 11.54). In the case of Banda Api, the 1988 eruption was preceded by three hours of strong earth tremors, and the post-eruption period (one week) was dominated by shallow explosion events (Casadevall et al. 1989) (fig. 11.55).
Evidence of earthquake damage is abundant on many reef slopes, especially in eastern Indonesia (fig. 11.56). Laminar and branching coral species are most susceptible to earthquake damage, however, large massive forms have been seen to be affected to varying degrees, as well (fig. 11.57). During volcanic eruptions massive colonies (e.g., Pontes, Diploastrea) can be displaced by the earthquake from the shallow reef slope to the base of the fore reef. Some of these displaced colonies manage to survive; others get buried by tephra, or by slumping of the reef talus during post-eruption tremors.

Figure 11.54. The 1988 eruption of Banda Api volcano. Seismic activity measured with Hosaka seismometer on Banda Api from April 1988 to May 1988. Plot of time versus s-p interval (seconds).
From Casadevall et al. 1989.
Figure 11.55. Plot of time versus magnitude for April and May 1988 using single-station, short-period Hosaka seismometer in Banda Api, Banda Islands. A) Shallow volcanic earthquakes with short period <4 sec. B) Deep volcanic earthquakes with short period >4 sec.
From Casadevall et al. 1989.

Figure 11.56. A large number of massive coral species, such as Porites lobata, were dislodged from the substrate during the 1988 volcanic eruption of Banda Api. The colony shown was displaced by the earthquakes from a depth of about 5 m to a depth of 25 m. Five years after the eruption the colony was still alive.
Photo by Tomas and Anmarie Tomascik.

Figure 11.57. Massive acroporids, such as Acropora palifera, are highly susceptible to earthquake damage. The earthquakes preceding and following the 1988 eruption of Gunung Api in the Banda Islands had a considerable effect on massive coral species. East coast of Banda Besar (Lonthor).
Photo by Tomas and Anmarie Tomascik.
Maumere Bay Earthquake
Earthquakes are obviously not always associated with nearby volcanic activity. Earthquakes triggered by tectonic forces can have a devastating effect on reefs. The massive earthquake that struck the eastern regions of Flores, and particularly the Maumere Bay area, on December 12,1992, had a measurable impact on the coastal and offshore coral reef communities as well as on the reef structure itself. This particular earthquake (6.5 Richter scale) also resulted in a tsunami that washed ashore at Maumere Bay and along most of the northeastern coastline, causing considerable devastation.
The reports from Maumere Bay following the December 1992 earthquake and tsunami reported widespread damage to human habitations as well as severe damage to coral reefs. Our survey of the area in November 1993 indicated that the structural damage was on a massive scale, however, after closer examination of numerous sites it became apparent that, while the earthquake did cause considerable structural damage to the reef base (fig. 11.58), the damage to coral communities was not consistent with the type of damage expected from an earthquake or a tsunami, which usually do not have much of an impact on deep-water benthic communities.
Based on our extensive survey of the area we came to the conclusion that the major damage to coral communities can be attributed to Cyclone "Lena", which affected the area between January 23-25,1993 (see box 11.2). Clear signs of cyclone damage, on a rather massive scale, were visible everywhere. Entire reef flats along the southern coastline of Pulau Besar, which were not affected by the tsunami, were pulverized and the rubble was transported considerable distances into the back-reef area, which clearly cannot be caused by an earthquake. At the base of the drop-offs, most of the rubble consisted of branching corals from the shallower reef flat and reef slope areas which were totally bare and pavement-like in character. While earthquake damage is mainly structural in nature, it significantly increases the vulnerability of coral communities to subsequent storm events. It is probable that the earthquake was to a large extent responsible for the severity of damage sustained during the subsequent cyclone. Numerous massive colonies dislocated from the substrate during the earthquake (fig. 11.59) were subsequently rolled over the reef by the force of the waves during the cyclone, thus pulverizing most shallow reefal areas.
Interestingly enough, in areas where large massive coral species were not in great abundance, structural damage to coral communities was not as extensive, and the recovery by November 1993 was substantial. Note that similar destruction of coral communities has not been reported from any of the adjacent areas that were also struck by the cyclone winds, but not by the earthquake. During our survey of the southeast Banda Sea (October 1993), we did not observe any storm damage in any of the areas that were most likely affected by Cyclone "Lena".

Figure 11.58. Earthquakes can cause considerable structural damage to coral reefs, especially along the seaward margins and steep drop-offs. Note the deep fissure that opened up at the seaward edge of a steep drop-off.
Photo courtesy of Sao Wisata Dive Resort, Maumere Bay, Flores.

Figure 11.59. During the devastating December 1992 earthquake in Maumere Bay, most large coral colonies were dislodged from the substrate and rolled over the reef during the January 1993 cyclone "Lena". Note the substantial regeneration (survival?) of tissue at the lower periphery of the Porites colony.
Photo by Tomas and Anmarie Tomascik.
TSUNAMIS
Generating Forces
The Japanese word tsunami literally means "long port wave" ("tsu" is port, "nami" is longwave) (Hadikusumah 1994). The original Japanese term has been accepted worldwide to classify ocean waves of exceptionally great wavelength caused mainly by seismic disturbances (e.g., submarine earthquakes) and/or volcanic activity. Tsunamis are often referred to as giant tidal waves, which is not correct, since they have nothing whatsoever to do with tidal-generating forces (i.e., gravitational fields of the Moon and Sun). Earthquakes are the main generating force of most tsunamis. There are a number of mechanisms by which earthquakes can generate tsunamis, but the two most important are associated with crustal displacement along submarine fault lines or massive submarine landslides on island shelves, both of which result in massive displacement of water. In addition, tsunamis are also caused by failures of the sea floor in the form of submarine slumping, generated by gravitational instability which may be triggered by an earthquake. However, eruptions of submarine volcanoes, while not as frequent as earthquakes, can generate tsunamis of greater magnitude than those generated by earthquakes. In general, any physical activity that will exert a significant force on the water column and cause a major displacement of its mass has the potential to generate a tsunami.
Physical Properties of Tsunamis
In the simplest terms, a tsunami is a progressive long wave, or gravity wave, which behaves as a shallow-water wave (i.e., its velocity depends on the depth of the ocean). As a result, tsunamis are influenced by the depth of the ocean and the physiography of the coastline. The characteristic features of tsunamis as they travel over open ocean are their exceptionally long wavelengths (i.e., >150 km), small wave heights (e.g., 0.5-1 m) and high velocities, that can reach up to 250 m.s-1 (900 km.hr-1). Thus, tsunamis traveling at the speed of commercial passenger jetliners can cross the oceans and wash ashore thousands of miles away from their place of origin, in a relatively short time. However, tsunamis usually achieve their greatest height, and thus their maximum destructive power, near their place of origin.
The destructive nature of the tsunami is manifested only upon reaching the coastline. In the open ocean, the passage of a tsunami with a wave height of 1 m and a wave length of 100 km would not be noticed on a ship. However, upon reaching a gently sloping coastline, the tsunami rapidly turns into a fast-moving wall of water, with an enormous amount of energy behind it. As the approaching tsunami encounters the bottom of the shelf, the wave velocity along the bottom of the shelf is reduced due to friction, relative to the crest, and refraction will cause the crest of the wave to align parallel to the coastline. However, since the velocity of the crest is not significantly diminished due to its long wavelength, and the amount of potential energy remains the same, the wave height begins to increase rapidly until it reaches its maximum height and breaks. The height of a tsunami (i.e., vertical distance between flat sea level and the crest) can vary from only a metre to more than 10 m. The destructive nature of a tsunami relates to its height and velocity, since most damage occurs from the direct physical impact of a rapidly moving wall of water against all objects in its path. Secondary effects relate to inundation of land and scouring (Birowo et al. 1983).
Tsunamis in Indonesia
As a result of its geological and tectonic setting as well as coastal physiography, the Indonesian Archipelago is one of the most tsunami-prone regions in the world (fig. 11.60). Since the devastating 1883 tsunami that was generated by the eruption of Krakatau, a total of 35 tsunamis were reported in Indonesia. Between 1960-1981 alone, six major tsunamis generated by offshore shallow earthquakes of more than 5.2 on the Richter scale claimed 1038 lives (Birowo et al. 1983). The 1979 tsunami that killed 700 residents of Lamalera, Lembata Island, was caused by a submarine volcanic eruption which occurred only a few kilometres offshore. Recent tsunamis in Maumere Bay (Flores, December 1992) and Banyuwangi (East Java, June 1994) claimed over 2000 lives (Okal 1993; Yeh 1993; Hadikusumah 1994; Helfinalis 1994).
Because of the frequency of shallow submarine earthquakes (70% of all earthquakes in Indonesia are submarine), which occur at relatively short distances offshore of most islands, tsunamis in Indonesia tend to be highly destructive when they do occur. Note that the destructive power of a tsunami is not necessarily correlated to the magnitude of the earthquake which may have caused it. Factors that determine the destructive potential of a tsunami are: 1) earthquake magnitude and depth of the epicenter; 2) area and speed of crustal displacement; 3) bathymetry in the path of the propagating tsunami; 4) direction of propagation; and 5) coastal physiography (Birowo et al. 1983). According to Birowo et al. (1983), historical records indicate that most tsunamis in Indonesia are of local origin, caused mainly by earthquakes, volcanic eruptions and landslides. Perhaps the largest tsunami ever to be recorded occurred following the 1883 eruption of Krakatau.
The Krakatau Tsunami
The Krakatau tsunami, which may have reached a height of 40 m at Merak, devastated the entire coastline of the Sunda Strait, and wreaked havoc along the west coast of Sumatra as it washed over the low-lying coastal plains (Birowo et al. 1983).
The damage from the Krakatau tsunami was excessive, with over 36,000 casualties and 250 villages and thousands of fishing boats destroyed. The velocity of the tsunami was estimated at over 600 km.hr", which did not leave much time to take refuge on higher ground. The tsunami propagated in all directions and was recorded by tide gauges in the western regions of the Indian Ocean at Cape of Good Hope, some 13,032 km away. Passing Sri Lanka, the Krakatau tsunami, still 1 m in height, claimed its last victim (Wilson 1992). To the east, the tsunami was recorded as a series of waves in Panama, over 20,600 km away (Birowo et al. 1983).
The now-famous "Cerita Beach" (cerita- story or an account) derived its name from this catastrophe, when thousands of bodies washed ashore along the entire length of the beach, victims of the tragic story of Krakatau (an account of an old fisherman).
The signs of the tsunami can still be seen today along the coastline of the Sunda Strait, where massive coral boulders were carried hundreds of metres inland. What type of damage the tsunami inflicted upon the coral communities of the coastal fringing reefs is not known. However, based on the size of some large Pontes spp. (3-4 m in diameter) found just north of Cerita Beach, it seems that many of the massive corals must have survived. With the exception of the 1983 survey by Sukarno and Suharsono (1983), this area has been generally neglected by the scientific community.
The Maumere Bay Tsunami
One of the most devastating tsunamis in recent times occurred in Flores on December 12, 1992. The tsunami followed a submarine earthquake with a magnitude of 6.5 on the Richter scale. The tsunamis that washed ashore at Maumere Bay reached heights of up to 4 m, while on Pulau Babi over 1000 inhabitants were killed by a 7.3 m tsunami that washed over the low-lying island. However, the tsunami reached its greatest height at Riangkrok (northeast Flores), where the tsunami left its deadly mark 26.0 m above sea level (Okal 1993; Yeh 1993). Huge coral boulders were dumped on the hillside, well above sea level, and 137 people lost their lives (Yeh 1993). What caused this exceptional height at Riangkrok is not known, however, massive subaerial landslides triggered by the earthquake may have been involved (Okal 1993).
Figure 11.60. Tsunami-affected regions (shaded) in Indonesia.
The earthquake and the subsequent tsunami that struck Maumere Bay and the northern coastline of Flores caused considerable damage and great suffering. Reports that arrived soon after the disaster indicated that the coral reefs sustained some serious damage. Because of the severity of damage to the local infrastructure by the earthquake, visits to the area were not possible prior to the tropical cyclone that affected die region on January 23,1993. As a result, the full impact of the earthquake and the tsunami cannot be ascertained, since the cyclone had considerably confounded the damage due to the earthquake and the tsunami. However, based on our November 1993 survey of the area, it seems that structural damage sustained during the earthquake was the main factor that made the coral community more susceptible to the cyclone. The damage by the tsunami is difficult to assess. However, it seems that it may have been minimal. For example, we found no evidence of large coral boulders on any of the reef flats affected by the tsunami. In comparison, huge coral boulders that were torn off the reef during the Krakatau tsunami litter the coastal plains along the Sunda Strait. The damage that we have observed on the reef flats and shallow reef slopes in Maumere Bay is consistent with damage reported from other parts of the world where cyclones are a frequent occurrence.
The Banyuwangi Tsunami
The latest deadly tsunami to strike Indonesia occurred on June 3, 1994, along the south coast of East Java, in the Banyuwangi district. The tsunami resulted in a loss of more than 200 lives, with 399 injured, as well as 1226 houses and 424 fishing boats destroyed (Hadikusumah 1994; Helfinalis 1994). The tsunami was generated by a shallow submarine earthquake (5.7-5.9 on the Richter scale) in the Indian Ocean (10°20' S and 113°20' E) just south of Java, at a depth of about 1500 m. The epicenter was located at a depth of 33 km. According to Helfinalis (1994), the earthquake may have originated as a result of sudden movement along the interacting Indo-Australian and Eurasian Plate margins. Faulting along the subduction zone is the most likely scenario that generated the Banyuwangi tsunami.
Based on the reconstruction of events from tide-gauge data at Cilacap and Banyuwangi, the tsunami propagating towards Banyuwangi reached a velocity of about 365 km.hr ", with a wavelength of 152 km and a period of 25 minutes (Hadikusumah 1994). The tsunami's greatest height of 4 m relative to local topography (about 7-9 m above mean sea level) was recorded at Pancer village (08°35'29.3" S and 113°59'43.2" E).
No reports are available on the damage to the coastal shallow-water marine communities. However, the coastal vegetation along many stretches of the coastline was severely damaged. At Pancer village, coral debris (large massive corals 27 x 24 cm) from a reef located about 150 m offshore was dumped beyond the village. The coastline in this area retreated by about 20-30 m as a result of the tsunami hitting the relatively gentle slope of the beach.
BIOLOGICAL PERTUBATIONS
Research into natural biological perturbations of coral reefs in Indonesia have thus far concentrated on the crown-of-thorns starfish, mainly in the Java Sea (Aziz 1977; Aziz and Sukarno 1977; Darsono 1988; Suharsono 1991; Darsono and Soekarno 1994). Coral predators are known to have a significant impact on reef structure and development (Glynn 1976). This is particularly evident in the Indo-Pacific, where outbreaks of the crown-of-thorns starfish, Acanthaster planci, have affected, and indeed decimated, many reefs (Birkeland 1982; Moran 1986). The Acanthaster phenomenon has not been as intensely studied in the Indian Ocean region as in the Pacific (e.g., Great Barrier Reef), however, crown-of-thorns starfish populations have been monitored in Indonesia, with numerous undocumented reports of outbreaks.
The gastropod Drupella rugosa has been reported to have damaged reefs in the Philippines and other regions of the Indo-Pacific (Boucher 1986; Black and Johnson 1994; Johnson and Cumming 1995). Sea urchins (particularly Echinometra mathaei, Diadema setosum and Tripneustes gratilla) and also Culcita are common reef inhabitants in the Indonesian region and, although not coral predators themselves, can cause significant damage if they occur in high numbers, especially following natural perturbations (Bak and van Eys 1975; Ogden and Lobel 1978; Sammarco 1982b; Lewis 1985; Glynn 1988). Echinoids graze on algal turf on coral rock and through erosion of the rock surface may weaken the reef structure, as has been recorded in a number of studies (Sammarco et al. 1987; Bak 1990).