Chapter Ten
INTRODUCTION
The Indonesian Archipelago is one of the most complex marine domains on the planet. The complexity of the Indonesian marine environment (i.e., physical, chemical and biological) is a reflection of its geologic history and atmospheric-oceanic interactions. Indonesian seas are among the most productive in the world, and support important coastal and offshore fisheries that are a significant source of protein for millions of coastal people throughout the archipelago. The high productivity of the intra-archipelagic seas is related mainly to high discharge rates of terrigenous material (e.g., nutrients and organic matter) from numerous rivers on Sumatra, Java, Kalimantan, Sulawesi and Irian Jaya, and to various types of geostrophic upwellings which are most pronounced in the eastern parts of the archipelago and along the south coast of Java, Bali and the Lesser Sunda Islands (Wetsteyn et al. 1990; Zijlstra et al. 1990). Significant local upwelling also occurs in numerous straits throughout the archipelago, where high-velocity currents cause vertical entrainment of deeper water masses. In addition, many coral reefs in areas with high tidal ranges generate their own daily upwelling events during flood tides by tidal suction (Wolanski 1992). Cold updrafts experienced by divers along steep reef slopes and drop-offs are a manifestation of this common "upwelling" phenomenon throughout the archipelago.
In the eastern Banda Sea, high nutrient concentrations in the 150-300 m layer occur throughout the year, supporting high primary production rates (e.g., 500 g C.m-2.yr-1) (Tijssen et al. 1990; Wetsteyn et al. 1990). In the Aru Basin, primary production rates during the upwelling season (i.e., Southeast Monsoon) were measured at 7-12 g C.m-2.d-1, which are among the highest recorded in tropical seas (Gieskes et al. 1990; Tijssen et al. 1990).
A generalization can be made that marine and coastal productivity of the intra-archipelagic seas in the western parts of the archipelago are influenced to a greater degree by terrestrial processes, while the high productivity in the eastern sectors is driven by upwellings. In this chapter we use coral reefs to illustrate how various environmental factors affect shallow-water coastal ecosystems.
TEMPERATURE
Temperature and Reefs
It was recognized early in the history of coral reef science that coral reef development was limited to tropical and subtropical regions, where surface seawater temperatures do not get much below 18°C (Dana 1843; Vaughan 1918, 1919; Molengraaff 1928; Yonge 1940). Veron and Minchin (1992) have conclusively demonstrated that functional coral reefs in Japan are restricted to latitudes where sea surface temperatures do not fall below the 18°C minimum for extended periods of time. In addition, they suggested that the development of coral reefs may not be limited by the low temperature tolerance of reef-building corals, but rather, restricted by light, macroalgae and other biotic and abiotic factors (Veron and Minchin 1992).
High-latitude reefs in the Indo-Pacific as well as in the Arabian and Atlantic regions puzzled many early scientists. The effects of temperature on the distribution of coral reefs were not always as clear-cut as of other environmental parameters, such as light (Vaughan 1919; Gardiner 1931; Achituv and Dubinsky 1990). However, Darwin (1842) was well aware of the modifying influence of the warm Gulf Stream, which supports extensive reef development, albeit with a depauperate coral diversity, on the Bermuda Platform in the North Atlantic (32° N).
The Bermuda Islands, in 32°15' N is the point furthest removed from the equator, in which they [i.e., coral reefs] appear to exist; and it has been suggested, that their extension so far northward in this instance is owing to the warmth of the Gulf Stream.—DARWIN 1842
The northern outpost of Atlantic reef development on the Bermuda Platform is indeed directly attributed to the overriding influence of the warm water masses associated with the Gulf Stream, which originates in the Gulf of Mexico. As a result the sea surface temperatures are not only within the thermal limits of reef-building corals, but are also favourable for biogenic reef accretion. However, seasonal fluctuations in sea surface temperatures do occur, and these are most likely responsible for the lower coral species diversity when compared to the western tropical Atlantic. Recent comparative growth-rate studies of hermatypic corals (colonial and solitary), from Bermuda and the tropical western Atlantic, demonstrated that coral growth rates decrease with increasing latitude, mainly as a function of lower mean sea surface temperatures characteristic of higher latitudes (Tomascik and Logan 1990; Logan and Tomascik 1991).
While there is a general consensus that temperature has played a significant role in the Recent (i.e., Holocene) worldwide distribution of coral reefs (Achituv and Dubinsky 1991), the mechanisms through which this influence is manifested remains a topic of numerous research efforts. Direct mortality associated with exposure to low temperature is an obvious mechanism. Early studies on the thermal tolerance of reef-building corals showed that they can tolerate a relatively wide range of temperatures, at least for short time periods (Mayor 1917; Edmonson 1928; Yonge and Nicholls 1931). Thus, some hermatypic species such as Pocillopora damicornis (Indo-Pacific) and Siderastrea radians (Atlantic) are able to survive for a few hours at 15°C (Vaughan and Wells 1943), while other species, such as Astrangia danae (Atlantic), seem to grow well at temperatures as low as 14.5°C (McCloskey 1970).
Since reef-building corals seem to have wider thermal limits than coral reefs, thus wider geographic boundaries (both latitudinal and longitudinal), it seems probable that the boundaries of coral reef distribution are linked to subtle temperature effects on coral calcification rates (Wells 1956; Gladfelter et al. 1978; Jokiel and Coles 1977), growth rates (Glynn and Stewart 1973), and most importantly, to ecological processes that have a direct bearing on the competitive fitness of the reef-builders (i.e., reef constructors) in high-latitude macroalgae-dominated environments (Crossland 1988; Hatcher 1991, 1993). Temperature has a direct effect on the rate of metabolic processes in all organisms, and it has been shown that, with an increase of 10°C, the metabolic activity of various organisms, measured by oxygen consumption, approximately doubles (Hoar 1966). Therefore, it seems likely that the sensitivity of coral species to temperatures below 18°C is inversely proportional to metabolic rates and growth rates. A few coral species can tolerate temperatures as low as 15°C, however, calcification rates are greatly reduced. At temperatures above 31°-32°C, metabolism increases to such rates that calcification and, therefore, growth may also be reduced. Since temperature directly affects the solubility of calcium carbonate (i.e., CaCO3 is more soluble in cold water), it follows that the calcification (i.e., deposition of CaCO3) in reef-building corals may be limited in cold water, restricting vigorous reef development to only tropical and subtropical seas. In these environments biological mediated precipitation of CaCO3 can proceed at a faster rate than physical, chemical and biological destruction (Barnes and Chalker 1990). However, earlier suggestions have recently been questioned that coral reefs were restricted latitudinally because of direct temperature effects on reproduction (Edmondson 1946; Jokiel and Guinther 1978; Kojis and Quinn 1984; Jokiel 1985), survival of coral planulae (Rosen 1975), or feeding rates (Edmondson 1928; van Woesik 1995; Veron 1995).
Comparative studies on high-latitude coral reef systems support the hypothesis that temperature plays a key role in the distribution of coral reefs, through direct and/or indirect effects on the structural and functional dynamics of biological communities (Crossland 1982; Wiebe et al. 1982; Johannes et al. 1983). Since coral reefs are products of biogenic processes, the most obvious being calcium carbonate deposition by hermatypic corals and other calcifying organisms, it follows that temperature regulation of calcification rates may be a key mechanism through which temperature effects are manifested in coral reef distribution patterns. The existence of high-latitude coral reefs can be directly attributed to the modifying effects of warm oceanic currents (e.g., Gulf Stream, Kuroshio Current, Leeuwin Current) that create favourable environmental conditions at high latitudes for coral growth (i.e., calcification and skeletal extension).
Temperature and Corals
The temperature story gets more complicated when we consider the individual coral species tolerance to temperature (Jokiel and Coles 1977), or the tolerance of different regional populations of the same species or genera. It has been stated that corals, and other reef-associated organisms, live at temperatures close to their upper thermal limits (Johannes 1975). It was therefore suggested that because of coral's relatively narrow temperature tolerance range (i.e., stenothermic), temperature increases of only a few degrees above ambient (~ 2°-3°C) can result in reduced growth rates or widespread mortalities in most coral species (Neudecker 1987; Jokiel and Coles 1990). Temperatures exceeding 33°C usually elicit a phenomenon called "coral bleaching", which is the expulsion of the symbiotic zooxanthellae from coral tissue by the coral animal. The presence of a large number of bleached (i.e., white in colour) coral colonies is a good indicator of possible environmental stress that may be caused either by natural processes (i.e., increased surface water temperatures during El Niño events) or by human impacts (i.e., thermal power plant effluents or other stressors). It should, however, be pointed out that most of the studies conducted on coral temperature tolerance are based on laboratory experiments.
Coles et al. (1976) demonstrated that the upper lethal temperature limits of tropical (i.e., Enewetak Atoll, Marshall Islands) corals are approximately 2°C higher than congeners from subtropical regions (i.e., Hawaii). If these results represent a general pattern, then we would expect corals from regions with higher ambient sea surface temperatures to have a tolerance for higher temperatures. If higher tolerance levels are linked to higher average ambient sea surface temperatures, as was demonstrated by Coles et al. (1976), then this type of adaptation should also occur on regional or even local scales. Field observations indicate that, indeed, some coral species are able to adapt to a relatively wide range of temperatures; thus we may see coral species existing in high-latitude reefs where average temperatures are at the lower limits, but are relatively stable throughout the year. On the opposite end of the spectrum we find corals living in areas with temperatures up to 42°C. Therefore, corals are able to acclimate to a wide range of local temperature regimes within the temperature tolerance range of the species over a short period of time. However, sudden and major deviations from the acclimated temperature regimes may trigger bleaching or be lethal.
Figure 10.1 illustrates four different sea surface temperature regimes from four different regions of the Indo-Pacific. The climate in the Marshall Islands and the Moluccas (i.e., Banda Sea) is clearly tropical, while Hawaii is considered by some to be subtropical (Coles et al. 1976). It is clear, however, that average monthly sea surface temperatures in the Marshall Islands do not fluctuate as widely as in the Moluccas. The difference in sea surface temperatures between these two regions is attributed to strong seasonal upwelling in the Banda Sea, which occurs during the Southeast Monsoon (May-September). Note that the upwelling in the Banda and Flores Seas during the Southeast Monsoon also has a considerable cooling effect on the sea surface temperatures in the Java Sea (fig. 10.1). The cooling effect in the Java Sea is probably related to the westward-flowing surface currents which bring in cool upwelled waters from the Banda Sea. The most intense upwelling occurs during July-August. Without the cooling effect, sea surface temperatures in the Java Sea would most likely be higher, probably comparable to sea surface temperatures in Jakarta Bay (fig. 10.2).
During the West Monsoon the temperatures in the Banda Sea are considerably higher than in the Marshall Islands. Sea surface temperatures as high as 32°C were measured on a number of occasions on shallow seaward reef slopes in the Banda Islands and southeast Seram (e.g., Koon Island). Across the archipelago in the shallow Java Sea, sea surface temperatures as high as 34°-36°C are common during low tides, especially in shallow lagoons and subtidal reef flats. However, these high temperatures are of relatively short duration, and with the incoming tide sea surface temperatures usually return to normal.
Gunung Api (Banda Api), in the Banda Islands, provides evidence that some coral species are able to recruit and survive for long periods in high-temperature environments. Gunung Api is an active volcano with considerable geothermal activity, which is indicated by the numerous shallow-water "hot vents" along the coastline. Submarine fissures from the 1988 lava flow on the northwest coast of Gunung Api are the main source of geothermal fluids. Most of the geothermal vents are intertidal; however, one subtidal vent was discovered at a depth of about 0.5 m below the lowest spring tide. Seawater temperatures within a few metres of the vent were 32°C (ambient 28.7°C), and the thermal surface plume was detected about 75 m down-current. The temperature of the geothermal fluid, a few centimetres above the vent, was 42°C. Growing within 2 m of the vent were numerous healthy colonies of Acropora florida, A. danai, Pocillopora damicornis and P. verrucosa. The temperature recorded was 34°C. Located at the immediate periphery were mature colonies of Pocillopora damicornis, P. verrucosa and Mycedium elephantotus. The temperature at the periphery of the vent was 37°C. The most surprising find (Tomascik and Mah, pers. obs.) was a juvenile Mycedium elephantotus (4.2 cm in maximum diameter), which was growing inside the vent. Another interesting observation was that the polyps of the juvenile colony were fully extended during daylight, while those of the larger colony, on the periphery of the vent, were not. Seawater temperature in the vent was recorded as 42°C on a number of visits to the site. These observations suggest that scleractinian corals may not be as stenothermic as was once believed. It seems that under natural conditions, scleractinian corals have a high degree of genotypic (i.e., relating to the genetic constitution of an individual or a group) and phenotypic (i.e., relating to the visible properties of an organism or a group) plasticity to tolerate a wide range of environmental conditions. Rapid colonization of the 1988 lava flow on Gunung Api as well as rapid coral skeletal growth rates (but not necessarily calcification rates) may be linked to higher temperatures associated with the geothermal activity (R. Ginsburg, pers. comm.).
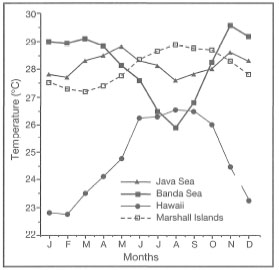
Figure 10.1. Comparison of average monthly sea surface temperatures in Java Sea, Banda Sea, Hawaii, and Marshall Islands.
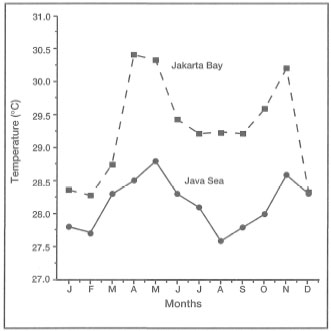
Figure 10.2. Annual sea surface temperature fluctuations in Jakarta Bay and Java Sea. Note the cooling effect of the Banda and Flores Seas because of upwelling during the Southeast Monsoon (May to September).
Coral Reefs and Upwelling
As discussed in the preceding section, temperature is now generally accepted to be the main factor determining the global distribution pattern of coral reefs (Veron 1995). Low temperature has been evoked as the main environmental factor responsible for the absence of coral reefs in areas of intense upwelling (Dana 1843), mainly along the western margins of continents. The west coast of South America (i.e., Galapagos Islands) and the west coast of Africa are the two most frequently cited examples. The west coast of Australia is an exception, since it is influenced by the warm Leeuwin Current, which has a modifying influence on the upwelling regime (Hatcher 1991). Considering the tropical nature of the Indonesian Archipelago, and that the archipelagic seas are the major route of the Indonesian Throughflow, which carries the warm western Pacific water (29°C), it seem unlikely that low temperatures are of any concern. Nevertheless, the eastern archipelagic seas are well-known for their intense upwelling during the Southeast Monsoon (Wyrtki 1961), and, therefore, the question of whether this may have an effect on the distribution of coral reefs within the archipelago needs to be addressed.
Based on the wide distribution of coral reefs, as well as non-reefal coral communities in the archipelago, it is clear that upwelling per se is not a significant factor determining coral reef distribution. In the eastern part of Indonesia, seasonal upwelling, driven by the Southeast Monsoon, occurs throughout much of the Flores and Banda Seas (Wyrtki 1961), two regions with the highest coral diversity in the archipelago (Best et al. 1989). In contrast, the eastern tropical Pacific, where upwelling systems occur along the western coasts of South and Central America, significantly reduces sea surface temperatures, thus determining regional distribution patterns of coral reefs. However, the seasonal upwelling system in eastern Indonesia does not result in a considerable drop of sea surface temperatures. Sea surface temperatures in upwelling regions are directly related to the intensity of the upwelling, which is mainly a function of wind strength and fetch. It is the intensity of the upwelling that determines the depth from which the water masses originate. The depth of the thermocline is another key factor that determines the sea surface temperatures in upwelling areas.
A generalization can be made that sea surface temperatures are much cooler in areas of strong upwelling than in areas of weak upwelling, since the water masses originate from greater depths in the former. Sea surface temperatures in the Banda Sea indicate that the upwelled water masses most likely originate from relatively shallow depths (100-200 m). For example, sea surface water temperatures in the Banda Sea are about 29°-30°C during the Northwest Monsoon (i.e., February/March), and 26°C during the Southeast Monsoon (i.e., August) (Zijlstra and Baars 1987; Boely et al. 1990; Ilahude et al. 1990; Zijlstra et al. 1990). Thus, sea surface temperatures during the strong Southeast Monsoon upwelling period are in fact at the optimal levels for reef development (Jokiel and Coles 1977). It has been shown that seasonal upwelling significantly enhances the primary and secondary productivity of surface waters through nutrient enrichment (Schalk 1987). Optimal temperatures, combined with abundance of plankton, suspended particulate matter and nutrients, may be one explanation why this region contains some of the most diverse coral reef communities in the archipelago. Habitat diversity may be another. Notably it is widely regarded that coral reefs have evolved in oligotrophic oceanic waters, which are often regarded as "nutrient deserts". However, this may be erroneous, since the area in the world that contains the highest coral diversity is also subjected to frequent upwelling and cannot be regarded as oligotrophic; in fact, organisms characteristic of oligotrophic and eutrophic environments are found living side by side (van Woesik, pers. comm.). It must be stressed that high-nutrient conditions that exist during the upwelling period cannot be equated with anthropogenic eutrophication, which has caused severe degradation of coral reefs worldwide (Bell 1992).
The seasonal upwelling also results in wider seasonal temperature fluctuations when compared to non-upwelling areas. Nevertheless, narrow sea surface temperature fluctuations are a characteristic feature of the archipelagic seas as illustrated by an example from the Flores and Banda Seas (fig. 10.3). It is clear that while the fluctuations are relatively small (26°-30°C), there are both latitudinal and longitudinal differences. Sea surface temperatures in the Flores Sea do not fluctuate as much as those in the Banda Sea, and the northern latitude stations show narrower ranges than the southern stations at the same longitude. These differences are linked with the more pronounced upwelling events in the Banda Sea as well as along the southern margin of eastern Indonesia. These minor shifts in sea surface temperatures most likely have no effect on coral communities; however, this is still pure speculation since comparative quantitative biological data are lacking.
The occurrence of localized upwelling events around individual islands has been corroborated by recent studies in the Banda Islands. In situ temperatures measured over fringing reefs during the upwelling events (range 24°C to 28°C), as well as temperature and nutrient depth profiles taken during the Snellius-II Expedition (Wetsteyn et al. 1990), indicate that the upwelling water masses which regularly spill over the fringing reefs in the Banda Islands originate from depths between 60 to 80 m. Intense localized upwelling, especially along vertical drop-offs, is also indicated by surface sea conditions (fig. 10.4).
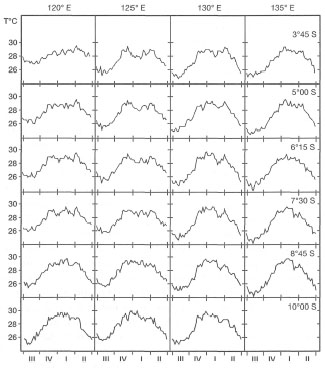
Figure 10.3. A) Sea surface temperatures (SST) in Banda Sea, illustrating progressive cooling of surface waters in easterly (left to right) and southerly (top to bottom) directions (i.e., towards the upwelling centres). Weekly data averaged over July 1981 to June 1985. Ill: July-September (Southeast Monsoon and upwelling); IV: October-December (transition); I: January-March (Northwest Monsoon); II: April-June (transition). B) Map of the four areas in Banda and Arafura Seas in A (above).
From Boely et a/. 1990. NJSR 25(4) Fig. 21 p.427.
Intense tidally-induced upwelling events (current velocities measured at 5-7 knots) may be a significant feature of Indonesian islands, and an important source of nutrients for the reef systems. Nutrient concentrations measured at these depths in the Banda Sea range between 0.13 - 0.53 μm and 0.9 - 7.8 μm of PO4 and NOs, respectively (Wetsteyn et al. 1990). High ambient nitrate concentrations may be one explanation for a recently demonstrated 8 N enrichment of Pontes spp. tissue samples collected from a fringing reef on Gunung Api (Heikoop, unpublished data). These values are similar to the 8 N values obtained from corals collected from a sewage-polluted area in the Maldives (Risk et al. 1993). The great diversity of coral reef communities in the Banda Islands suggests that seasonal monsoonally-induced upwelling, as well as the daily tidally-induced upwelling, may actually promote the productivity of coral reef communities, and thus reef development.
High Temperatures
High sea surface temperatures are not yet likely to be a significant factor in the regional distribution of coral reefs in the archipelago, however, they may have a significant local effect on community structure and reef zonation. High temperatures have been linked with reef-bleaching events (zooxanthellae expulsion) which resulted in extensive coral mortalities, thus affecting zonation patterns within reef systems. However, because of turbulent mixing associated with strong tidal currents, a dominant feature of the archipelagic seas, high sea surface temperatures, in general, do not seem to be a major factor in reef distribution patterns.
The 1983 ENSO-associated mass-bleaching event in the Java Sea has been attributed to a significant temperature rise; however, it occurred in a relatively isolated area and under a specific set of environmental conditions. In addition, all impacted reefs during the 1983 ENSO event were in areas of intensive development and under heavy exploitation pressure. For example, coral reefs in Kepulauan Seribu are being subjected to a variety of anthropogenic impacts (e.g., eutrophication, siltation, etc.) which may have made them more susceptible to natural disturbances. In other regions of the archipelago, temperatures as high as 36°C have been observed to cause no visible damage to corals (fig. 10.5), and healthy corals have been found to survive periodic exposures to temperatures as high as 42°C (fig. 10.6). Based on these observations, it may be concluded that the general coral reef distribution patterns within the archipelago are not temperature-related.
SALINITY
Definitions
In the simplest terms, salinity is the measure of the sum of all inorganic salts dissolved in seawater. The average concentration of dissolved salts in the world's oceans is roughly 3.5% by weight (i.e., 35 psu - physical salinity units, which is equivalent to ppt and %o) (Dietrich et al. 1980). Salinity in the oceans varies greatly among regions as well as with depth. Salinity is a conservative property of seawater, since changing the concentration of major salts (i.e., dissolved ions such as Cl- and Na+, etc.) will not alter their ratios (i.e., relative proportions), which remain constant irrespective of salinity changes. With the exception of the Red Sea and the Arabian region, oceanic sea surface salinities are highest in the Atlantic Ocean. Salinity changes have a profound effect on the physical properties of seawater, since salinity causes variations in specific gravity, and therefore density, which has a direct effect on the movement of water masses and circulation patterns.


Figure 10.4. A) Tidally-induced upwelling eddy along a reef drop-off at Koon Island, southeast Seram (eddy temperature 23°C; reefal sea surface temperature 28.5°C). B) Current-induced upwelling eddy in Boleng Strait, between Adonara, Solor and Lembata Islands, East Nusa Tenggara (eddy temperature 22°C; normal sea surface temperature 29.4°C).
Photos by Tomas and Anmarie Tomascik.
Within the Indonesian Archipelago, sea surface salinities vary greatly along both spatial and temporal scales. Different climatic conditions throughout the archipelago, large islands with high volumes of river runoff, various geological barriers (i.e., island arcs), and sea surface circulation patterns all contribute to significant salinity differences among the various archipelagic seas. It is well-known that chemical properties of seawater have a direct effect on the basic metabolism (osmoregulation and other physiological processes) of marine organisms, including corals, and therefore, salinity has a significant influence on their distribution, including the distribution of coral reefs. For many reef organisms, and especially corals, salinity determines survival, spawning areas, optimum growth conditions, and movements during life history changes. In the following discussion on the effects of salinity, most experimental studies to date were on mature coral colonies. Recent experiments in Guam and Okinawa have clearly demonstrated that a 20% reduction in seawater salinity causes an 86% reduction in fertilization success in corals (Richmond 1993). The question here is whether regional and local salinity differences, within the archipelagic seas, are sufficient to affect the distribution of coral reef communities and coral reef development in general.

Figure 10.5. Sangalaki Island, Berau Islands, East Kalimantan. The large shelf reef with a densely vegetated cay is located about 60 km offshore. Sea surface temperatures on the intertidal reef flat reach 36°C at low tides (tidal range 2.75 m). Intrusions of upwelled water masses are common.
Photo by Tomas and Anmarie Tomascik.
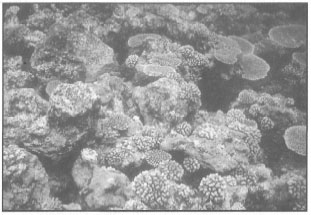
Figure 10.6. Banda Api shallow-water coral community subjected to hydrothermal fluids and daily heating at low tide (tidal range 2.5 m). Sea surface temperatures as high as 38°C were recorded, however, coral bleaching is not a major problem.
Photo by Tomas and Anmarie Tomascik.
Salinity and Global Reef Distribution
The fact that coral reefs were absent from all tropical regions heavily influenced by river runoff is a well-known fact since Darwin (1842). Salinity, at first, seems a simple explanation, since extensive coral reef development has never been reported in any region where the average sea surface salinities are below approximately 29 psu. This includes all regions affected by the world's major river systems (table 10.1).
For example, in the Atlantic, major coral development occurs in regions where sea surface salinities range between 34-36 psu (i.e., Caribbean). The Amazon River, with an annual runoff of about 5.7 x 1012 m3, has a far-reaching influence on the distribution of coral reefs along the northwest coast of Brazil, and perhaps also influences the general nutrient dynamics of the eastern Caribbean Sea. As a result of the North Equatorial Current, the Amazon low-salinity plume travels in a northwesterly direction, effectively inhibiting major reef development until the Lesser Antilles (i.e., Trinidad and Tobago), about 2100 km away. These low-salinity eddies, which are distinct water masses, are seasonally detectable as far north as Barbados, 2400 km from the Amazon delta. The northwest coastline of Brazil is, however, fringed by extensive seagrass beds and mangal forests, while the Brazilian Abrolhos to the south support flourishing coral reef communities. Recent questions concerning the regional effects of the Amazon River plume on Caribbean reef nutrient dynamics has attracted new research efforts in the region. Coral reef regions of the Pacific Ocean and the central Indian Ocean (e.g., Maldives) maybe considered as relatively unaffected by continental processes (with the exception of some high oceanic islands such as Hawaii, Kosrae, etc.). Sea surface salinities within the subtropical and tropical regions of the Pacific Ocean range between 35-36 psu, with the exception of a small region extending from about 90° W to 130° W (between 10° N20° N), where the average salinities are about 34 psu (fig. 10.7)
In the Southeast Asian region, coral reefs occupy a variety of environments, ranging from the turbid shallow continental seas (e.g., Gulf of Thailand, Java Sea, Arafura Sea) to deep oceanic basins where atolls and high-island reefs (i.e., fringing and barrier type) developed in clear oceanic waters free of terrestrial influence (e.g. Andaman Sea, Sulawesi Sea, Flores Sea, Banda Sea). The western extent of the Southeast Asian coral reef distribution can be delineated by the Andaman and Nicobar Islands, located to the north-northwest of Sumatra. The Andaman-Nicobar reefs developed on a submarine ridge that was formed at the northern extension of the Sunda Trench subduction system. In fact, the Andaman-Nicobar Ridge is a continuation of the non-volcanic Mantawai-Nias Outer Arc Ridge, which is part of a continuous subduction system along the Sumatra Trench (Hutchison 1989). These reef systems are sufficiently distant from the Ganges-Brahmaputra and Irrawaddy River systems, whose combined annual runoff volume is about 1.1 x 1012m3. As would be expected, the Ganges-Brahmaputra delta (the largest subaerial delta in the world) supports extensive mangal forests. In the Gulf of Thailand, river runoff has a significant effect on the distribution of coral reefs. Low sea surface salinities (20 psu) in the northern part of the gulf can extend up to 70-80 km seaward from the Chao Phraya River delta (Piyakarnchana 1981), thus limiting coral development to the more southern regions of the gulf.
Table 10.1. Mean annual flow rates of the Mahakam and Solo Rivers, and some of the major world river systems.
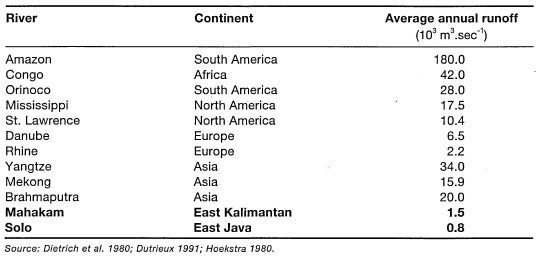
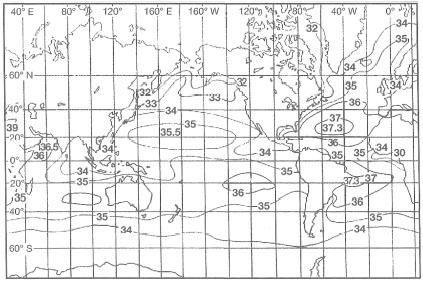
Figure 10.7. Distribution of sea surface salinities throughout the world's oceans.
From Open University 1991.
Upon close inspection, however, the salinity control hypothesis becomes problematic, especially in the Southeast Asian region. River runoff not only dilutes the salinity of the receiving coastal waters, thus subjecting marine organisms to osmotic stress, but all major tropical river systems carry a heavy load of suspended sediments that may be transported long distances from the river, particularly during storm events. Thus, a clear separation between salinity, turbidity and sedimentation as the major causative factors in reef distribution is extremely difficult to demonstrate (Fagerstrom 1987). Interestingly enough, Darwin (1842) was of the opinion that, while reduced salinities unquestionably have a negative impact on corals (i.e., physiological stress), the main factor responsible for determining the geomorphology and distribution of inshore coral reefs, at least on a local scale, is the deposition of fluvial sediments (i.e., sedimentation). This process seems to be the main factor determining the distribution of many inshore reefs within the Indonesian Archipelago and Southeast Asia in general.
No doubt brackish water would prevent or retard the growth of coral; but I believe that the mud and sand, which is deposited, even by rivulets when flooded, is a much more efficient check.—DARWIN 1842
In some arid regions of the world (e.g., Bahrain), reef-building coral communities are known to flourish in environments where salinities seldom drop below 42 psu. Extensive reef development in the Red Sea and the Arabian region supports this fact. For example, Porites nodifera-dominated communities form patch reefs in environments where salinities range between 43-45 psu (Sheppard 1988). Short-term exposure to high salinities have been reported earlier, but Sheppard (1988) found that some coral species (e.g., Porites nodifera, Siderastrea savignyana and Cyphastrea microphthalma) can survive in salinities as high as 50 psu. However, it is evident that only marginal coral reef development occurs in these high-salinity environments, and the coral communities are dominated by only a few tolerant coral species (28 species in Bahrain) (Sheppard 1988).
Based on the wealth of information from the Red Sea and Arabian regions, it seems that scleractinian corals have a remarkable physiological tolerance (i.e., acclimatization ability) to high saline conditions (Coles 1993). However, coral survival or reef development at salinities similar to or below the normal minimum have not been demonstrated (i.e., 15 psu below the lower normal value of 30 psu). Vaughan (1919) found that some Caribbean coral species survived in salinities of 28 psu for 24 hours, while other studies indicated greater sensitivity. Note that coral tolerance to low salinities, under laboratory conditions, is not indicative of their ability to survive under these conditions as viable populations, nor is it indicative of reef-forming capability. What the laboratory experiments have shown is that corals are able to tolerate above- or below-normal salinities, for certain periods, which is not surprising since on a global scale coral reefs encounter a wide range of salinities (Stoddart 1969).
Coles and Jokiel (1992) have reviewed the available salinity data from numerous coral reef studies around the world, and came to the conclusion that: "corals and other reef organisms can live in normal salinities as low as 25%o and as high as 45 %o". However, out of the 38 studies cited in the review, only six studies reported salinities below 30 psu, the lowest value of 25 psu being from the Gulf of Thailand. Closer inspection of the Thailand data (Piyakarnchana 1981) reveals that the reefs in the northern regions of the gulf were outside the 25 psu isohaline. The fact that low salinity water is usually restricted to the 1 to 3-m-deep surface layer was not considered. The reevaluation of the available data (Coles and Jokiel 1992) suggests that there is no evidence to support the view that vigorous coral reef development occurs at salinities below about 29 psu. It is also clear that reef-building corals are not as stenohaline as was once assumed. In fact, many coral species have a wider salinity tolerance range than other reef-associated organisms.
Based on the worldwide distribution of coral reefs, and experimental data, it seems that reef-building corals are able to tolerate salinities of about 25 psu for brief periods. This suggests that periodic exposures to low salinities, which may, for example, occur during tropical storms, will not inhibit reef development. It may be concluded that reef-building corals and coral-reef-associated organisms generally (there are exceptions) flourish in regions where average sea surface salinities are maintained between 30 to 36 psu, and therefore, coral-dominated "reef-building" may be loosely defined as a stenohaline process.
Salinity and Indonesian Reef Distribution
Sea surface salinities in the archipelagic seas are influenced by a number of environmental factors (e.g., runoff, precipitation, evaporation, surface current patterns). The most obvious, and the most easily interpretable, is the effect of river runoff during the Northwest Monsoon. Heavy rainfall during the wet season is often associated with floods and significant fluctuations in river discharge volumes. For example, during the Northwest Monsoon (November-January) the daily flow rates of the Solo River, with a catchment area of about 16,000 km2, may fluctuate between 300 and 1800 m3.sec4, while during the Southeast Monsoon the daily flow rates fluctuate between 80-250 m3.sec-1 (Hoekstra 1989; Hoekstra et al. 1989). Just to the south of the Solo River delta, the Brantas River discharges an additional 600-1200 m.sec-1 of runoff during the wet season (Heokstra et al. 1989), while the combined volume of these two major Javanese rivers (maximum wet season combined flow of 3000 m3.sec-1) is considerably less when compared to the flow rates of some of the other major rivers in Southeast Asia (e.g., 15,900 m.sec-1 for the Mekong River). These rivers nevertheless have a significant impact on the coastal geomorphology and the distribution of shallow-water coastal ecosystems along the northeast coast of Java and Madura. Since the coastal surface currents during the Northwest Monsoon are dominated by a net eastward flow, the Solo-Brantas River runoff may be a key factor responsible for the absence of major reef development along the north coast of Madura. Significant reef development does not occur until about 130 km to the east of the Solo-Brantas deltas, along the south coast of Madura (e.g., Pulau Giligilingan). However, marginal fringing reefs occur along the south coast of Madura, about 40 km east of the Solo-Brantas deltas (e.g., Labuan). The nearest reefs to the Solo delta in the east Java Sea are the fringing and patch reefs around Pulau Bawen, about 130 km to the north. However, it is suggested that the general absence of coral reefs in this region of the east Java Sea is probably associated mainly with the lack of suitable substrate, rather than with the effects of the Solo-Brantas River runoff. This is an interesting area of research in an accessible area.
As expected, there seems to be a general absence of coral reefs throughout the archipelago in the immediate vicinity of major rivers. However, closer inspection reveals that the effects of low salinities may be confounded by other environmental factors, such as reduced light levels associated with increased turbidity or sedimentation, as well as possible interactions of these parameters. Heavy runoff from the major rivers on Java Island (e.g., Solo River) during the Northwest Monsoon is most likely the main factor responsible for the absence of reefs along much of its northern coastline. To what extent salinity influences this outcome is difficult to determine, since clear-water, low-salinity, tropical coastal environments have not been studied. Lack of suitable substrate, high turbidity and/or heavy sedimentation along much of the northern coastline may be the overriding factors causing a dearth of reef development. There are a number of patch reefs situated off the north coast of Java (e.g., Gosong Tengah, Karang Rakit, Gosong Pamanukan, etc.) where the presence of suitable substrate has allowed coral reef development. These areas were originally far enough from the influence of natural river runoff, and as a result, coral communities became established and reefs were formed. Hydrological conditions along much of Java's north coast have changed in recent years (Verstappen 1953), mainly as a result of major changes in land drainage patterns. These changes in coastal hydrodynamics are associated with large-scale conversion of coastal plains into agricultural land (i.e., wetland paddy fields) as well as deforestation and land conversion of upper water catchment areas. Both of these practices have resulted in increased freshwater runoff and a high load of suspended solids carried down the rivers during the wet monsoon.
As a result of macro-scale weather patterns associated with the monsoonal system, as well as the high mountain ranges, the Indonesian Archipelago is not climatically homogeneous. When viewed from a regional level, there is a distinct annual rainfall pattern present. Figure 10.8 illustrates that the west (e.g., Sumatra, Kalimantan, Java) and east (e.g., Irian Jaya) regions of the archipelago receive considerably more rainfall on an annual basis than the central regions (e.g., Lesser Sunda Islands, Moluccas). The lowest annual rainfall occurs in the Lesser Sunda Islands. The central region of the archipelago, with the Philippines, was a known centre of generic coral diversity (Best et al. 1989; Veron 1986, 1995). However, Hoeksema (see box 7.2; chapter 7) recently expanded the generic diversity of this region; thus for the time being it may be considered as a high-diversity sub-centre within the Indonesian-Philippine Province.
The differences in rainfall between the various regions of the archipelago, however, are not only of quantity. With regards to coral reef development and coral community structure, the seasonal patterns of rainfall may be more significant than the average annual values. It is often stated that the archipelago is under the influence of a monsoonal climate, and therefore, marked seasonality in rainfall should be expected. However, as is clearly demonstrated in figure 10.9, there are distinct differences not only in the amount of rainfall, but also in the seasonality.
For example, average monthly rainfall in Fak-Fak, located on the Onin Peninsula in Irian Jaya, ranges between 250-400 mm without any pronounced seasonal fluctuations. The seasonal weather pattern in this area can best be described as either wet or very wet. Similar conditions occur in Tarakan, East Kalimantan, where the average monthly rainfall varies between 250-300 mm throughout the year. Strong dry-wet seasonality is, however, characteristic of much of the archipelago, especially in the southeastern regions (i.e., Flores, Timor, Komodo, etc.). Smaller areas within each region can also exhibit marked differences as a result of microclimate variability. In these areas the climate is influenced mainly by local physiography. Local differences in seasonal patterns in average monthly rainfall are illustrated in figure 10.10. These regional differences in rainfall patterns have a measurable effect on the hydrology of the coastal area, and therefore, on the distribution of coral reefs. The coastline of most eastern islands is fringed by extensive fringing reefs. For example, the island of Yamdena, in the Tanimbar Islands, has a fringing reef that runs virtually uninterrupted along the entire length (i.e., c. 375 km) of the coastline, except where breached by rivers and rivulets. Some of the most diverse coral communities are found in the drier eastern regions of the archipelago, characterized by strong seasonality in rainfall and low volume of land runoff. However, this is not to say that diverse coral communities, or extensive coral reef systems, have not developed in areas of continually high rainfall. The extensive barrier reef system in the Berau Islands, in East Kalimantan, illustrates that major coral reef development can occur in relatively turbid, shallow-water coastal areas in close vicinity to major river systems (see fig. 10.5).
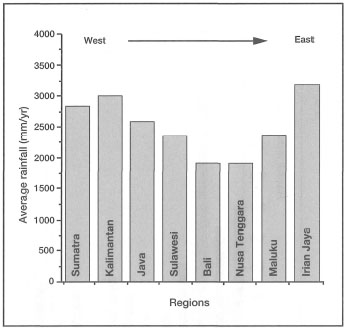
Figure 10.8. Average yearly rainfall (mm) for the main physiographic regions of the Indonesian Archipelago.
Source: RePPProT 1990.
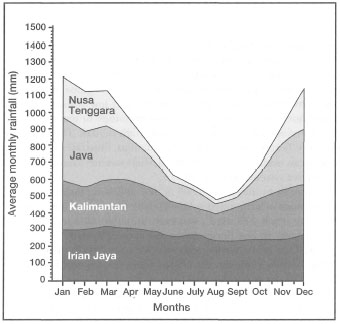
Figure 10.9. Average monthly rainfall (mm) for Kalimantan, Java, Nusa Tenggara and Irian Jaya, demonstrating different climatic conditions. Continually wet climate - Irian Jaya; Slight seasonality in rainfall - Kalimantan; Dry and wet monsoons Java; Dry and arid conditions - Lesser Sunda Islands.
Source: RePPProT 1990.
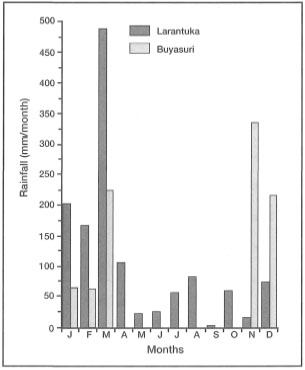
Figure 10.10. Local rainfall patterns. Comparison of average monthly rainfall between Larantuka, east Flores and Buyasuri, Lembata Island. The two districts belong to the East Flores Regency, East Nusa Tenggara. The distance between the two locations is 60-70 km. Rainfall data from 1990.
Source: Pemerintah Kabupaten DATI II, Flores Timur 1992.
Flow rates in the Berau River may peak at 500 m3.sec-1 following heavy rains, and carry a considerable amount of suspended solids. During December SPM concentrations in the delta may reach 175 mg.1-1, however, available data indicate that average SPM concentrations in the Berau River are about 80 mg.1-1 (Hatfindo, pers. comm.). The fluvial fan of the Berau delta extends some 20 km offshore. To the north of the Berau Islands, however, the coastal water quality rapidly deteriorates from the influence of the Bulungan and Sesayap Rivers. Karang Baliktaba is the last coral outpost at the northern fringe of the Berau Barrier Reef (fig. 10.11).
The Baliktaba Reef is located about 30 km to the southeast of the Bulungan River delta, just at the outer fringes of the turbidity zone associated with the Bulungan River plume. The coastline between Karang Baliktaba and Tarakan is fringed by extensive mangrove and Nypa fruticans forests. However, high turbidity of the coastal waters loaded with fluvial sediments seems to be the main factor responsible for the absence of coral reefs from this area, since surface salinities are above 30 psu. The lack of a suitable substrate also may be important, considering that the shallow-water habitats seaward of the mangrove belt are mainly mud flats. Shallow-water and high-velocity tidal currents rapidly mix the river water with coastal water masses, and help to maintain high concentrations of fluvial sediments in suspension for long distances. The fluvial fan of the Bulungan River extends over 20 km offshore, where extensive mud banks have been formed. The complexity of the abiotic factors makes it impossible to ascertain what role, if any, salinity plays in the distribution of coral reefs in this region.
Figure 10.11. The Baliktaba reef complex is the last coral outpost at the northern boundary of the Berau Barrier Reef (see fig. 12.4, chapter.12). The reef rises from a depth of about 50 m and is surrounded by a mixture of fluvial and bioclastic sediments. A) The intertidal and shallow sub-tidal habitats are dominated by acroporids, supporting a diverse reef fish fauna. B) A more diverse coral community is found on the deeper reef slope. The reef extends to a depth of about 15-20 m.
Photos by Tomas and Anmarie Tomascik.
Effects of Low Salinities
Sea surface salinity fluctuations, associated with river runoff and/or heavy rainfall, may be a key factor determining local (i.e., within reef) zonation patterns, especially in coastal areas with large tidal ranges. Coral reefs located in close proximity to large rivers (e.g., Berau River, East Kalimantan) exhibit a marked transition from low-diversity reef flat communities which are dominated by a few hardy coral species (e.g., Pontes lobata, P. lutea, P. cylindrica, Montipora turtlensis, M. mollis, M. hispida, Goniastrea favulus, G. aspera, Goniopora djiboutiensis, G. stokesi, G. lobata) that are generally tolerant of high turbidity and lower salinities, to high-diversity reef slopes that support luxuriant coral growth further from the river (fig. 10.12). In contrast, reef flats of oceanic platform reefs (e.g., Nil Desperandun, Banda Sea) are high-energy coral communities, dominated mainly by luxuriant assemblages of acroporids (e.g., Acroporapalifera, A. robusta, A. danai, A. austera, A. cytherea, etc.) as well as other coral groups (fig. 10.13). Note, however, that oceanographic conditions (i.e., currents and waves) determine zonation patterns of oceanic coral reef communities removed from the influence of terrestrial processes.

Figure 10.12. The Tanjung Batu Reef is located about 1.5 km offshore, at the seaward limit of the Berau River delta (see fig. 12.4 for location). The apparently lagoonless shelf reef, with an area of about 30 km2, is cut by numerous narrow channels. A) Mainly as a result of its sheltered location, the intertidal reef flat coral community, on Tanjung Batu Reef, has a distinct lagoonal character (tidal range 3 m). The Berau River plume frequently impinges on the reef; however, as a result of strong southward currents the reef is maintained relatively sediment-free. Note the presence of Pontes microatolls. B) The deeper reef slope (4-5 m) of Tanjung Batu Reef has noticeably higher coral diversity and coral cover, corresponding to increased water clarity.
Photos by Tomas and Anmarie Tomascik.
Based on observations from East Kalimantan, the north coast of Java, and the southwest coast of Irian Jaya (e.g., Bintuni Bay, etc.) we arrive at the expected conclusion that coral reefs are not found along coastlines characterized by predominantly estuarine conditions (e.g., widely fluctuating salinities and high turbidity), and extensive tidal range. Unfortunately, the role of salinity in this well-known dictum is not very clear. However, it is well recognized that substantial damage and mortality can occur with flood waters that significantly reduce sea surface salinities below ambient. For example, Glazebrook and van Woesik (1993) and van Woesik et al. (1995) documented that corals suffered about 85% mortality when floodwater reduced salinities to 8%o following a major storm. Dead corals were rapidly overgrown by turf algae two weeks after the flood event. Coral mortality was restricted to depths less than 1.3 m, however, reef-bleaching was observed at greater depths. Tissue samples collected from corals subjected to low-salinity stress indicated hypertrophy, hyperplasia and lysis of the epidermis as well as degenerative changes in the endodermis, which sometimes extended to necrosis (Glazebrook and van Woesik 1993; van Woesik et al. 1995). In addition, bacterial emboli were present in the subepidermal tissue layer (van Woesik et al. 1995).

Figure 10.13. Subtidal reef flat on Nil Desperandun, a submerged oceanic platform reef in the Banda Sea. The subtidal reef flat is dominated by a diverse Acropora community, predominantly by Acropora palifera and A. robusta. Tidal currents sweeping over the reef were recorded at about 4.5 m.sec-1 (8.7 knots).
Photo by Tomas and Anmarie Tomascik.
Sea surface salinities as low as 25 psu (found in the Gulf of Thailand) may not be uncommon in many coral reef areas, and have indeed been observed in Jakarta Bay during the Northwest Monsoon (Tomascik, pers. obs.) as well as during the regular oceanographic cruises of the R.V. Samudra. Ilahude and Liasaputra (1980) reviewed oceanographic reports from Jakarta Bay and their results clearly demonstrate a sea surface salinity gradient extending from the Citarum and Bekasi Rivers in a northwesterly direction towards Kepulauan Seribu (fig. 10.14).
While sea surface salinity data are useful in helping to delineate the horizontal boundaries of river plumes, depth profiles should accompany the salinity data to obtain the complete picture. Table 10.2 demonstrates that low sea surface salinities do not necessarily extend into the lower water column (i.e., freshwater, being less dense than seawater, floats on top), and therefore, may not directly impact upon deeper coral reef communities. Similar observations were made by Kastoro et al. (1989) just offshore of the Solo and Brantas River deltas in East Java, during the wet and dry seasons. The combined discharge volume of both rivers is considerable during the wet season, but the salinity effect is rather localized due to rapid mixing with seawater. During the rainy season, sea surface salinities 1 km offshore (depth 0.3-1 m) ranged between 25-28 psu. At 2 km offshore (depth 1-2 m) salinities increase to 30-31 psu, and 4 km offshore (depths 3.5-9 m) salinities ranged between 30-33 psu.
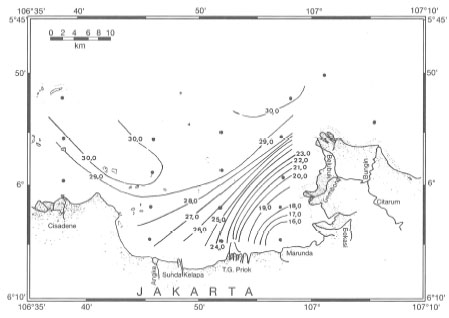
Figure 10.14. Normal sea surface salinities in Jakarta Bay during the Northwest Monsoon (January), characterized by heavy rainfall and river runoff.
From Ilahude and Liasaputra 1980.
Table 10.2. Salinity depth profile for Stations 44 and 45 in Natuna Sea, Oceanographical Cruise Report No. 3, R.V. Samudra, February 12 - March 5, 1971. Profiles at Stations No. 44 and 45 taken on February 25, 1971. No. 44: 02°14'00" N, 110°18'00" E' No. 45: 02°32'40" N, 109°56'10" E.
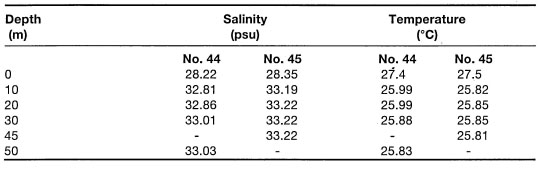
Note, that the coral reefs of Jakarta Bay were not long ago flourishing systems, which apparently evolved in relatively close proximity to major rivers (Verwey 1930a; Umbgrove 1939a). Low sea surface salinities such as those found in Jakarta , Bay require careful interpretation, since low-salinity water masses seem to be restricted mainly to the surface layer and generally do not extend below 3 m depth. This seems to be the case for Jakarta Bay, where a low-salinity surface layer (24.5 psu) was observed to extend to a depth of 2 m below the surface, thus not in direct contact with the depauperate coral community 5 m below. However, all rivers running into the Java Sea are heavily laden with sediments, and therefore, it is more than likely that significant reduction of light levels and sedimentation are the main impacts associated with low-salinity surface water masses. The effects of light, turbidity and sedimentation are discussed in the next section.
Berau Islands: Case Study
The influence of river runoff, and the difficulty in determining the effects of salinity, can be illustrated by an example from Karang Buliulin, which is located at the southern extension of the Berau Barrier Reef system, about 40 km downstream of the Berau River delta (see fig. 12.4). The intertidal reef flat, about 35 km in area, is under the influence of a strong diurnal tidal regime (F=0.23), with a maximum tidal range of about 2.8 m (fig 10.15).
The reef flat has strong 'lagoonal' attributes in terms of sediment characteristics and community structure. There are visually striking differences in terms of coral community structure and coral cover between the western and the eastern reef slopes (fig. 10.16). This difference may be attributed to the influence of the Berau River, whose sediment-laden plume impacts first on the west slope of the reef. The sediments at the bottom of the reef slope, along the west boundary of the reef, contain a higher fluvial fraction when compared to the more bioclastic sediments found along the eastern boundary of the reef.
In essence, the extensive intertidal reef flat of Buliulin Reef (4 km wide and 11 km long) acts as a "block", since water quality (in terms of turbidity) between the west and east regions of the reef are very different. Horizontal underwater visibility (at a depth of 5 m) on the west slopes was 5 m, while on the east coast the horizontal visibility extended to about 15 m. This area offers interesting research opportunities.
Salinity measurements demonstrated that localized upwelling of deeper water masses during the flood tide modify the effect of low-salinity eddies originating from the Berau River plume. At the end of the Northwest Monsoon, a large low alinity eddy (28 psu at 0.5 m) was detected over the entire reef area (Tomascik and Mah, pers. obs.). At high spring tide, the reef flat is about 2.5 m below sea level. Salinity at the bottom was 33 psu, thus the surface low-salinity plume was restricted to the top 1.5 m depth. During the flood tide, the tidal currents are flowing in a predominantly northwesterly direction with a velocity of about 1 m.sec-1. Sea surface temperatures measured over the reef flat (about 50 m from the outer reef slope) averaged about 30°C, while the bottom (2.5 m depth) reef flat temperatures and salinities were 28°C and 33 psu, respectively. The sharp temperature and salinity differences between the surface and bottom layers is related to localized upwelling generated by flood-tide entrainment of deeper water masses along the seaward edge of the reef (maximum depth 60 m) (fig. 10.17), known as the Bernouilli effect or tidal suction (Thompson and Golding 1981; Thompson and Wolanski 1984). The upwelling of more saline and cooler waters provides an effective barrier against the influence of low-salinity surface water masses, especially if this phenomenon is cyclic (i.e., tidal).
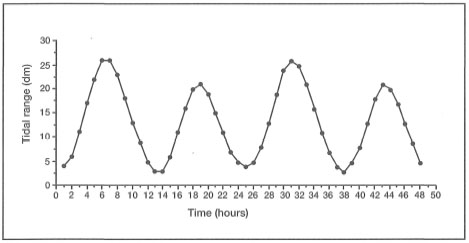
Figure 10.15. An example of a semidiurnal tidal cycle (F = 0.23) over a two-day period in the Berau Islands. Maximum amplitude is about 2.8 m.
Source: Daftar Pasang Surut 1994.

Figure 10.16, A) Reef slope on the west side of Karang Buliulin, Berau Islands, East Kalimantan. The west side of the large offshore patch reef is influenced by river runoff. B) Reef slope on the east side of Karang Buliulin. Note higher diversity, cover and improved water quality. Photos A and B were taken on the same day, within one hour.
Photos by Tomas and Anmarie Tomascik.
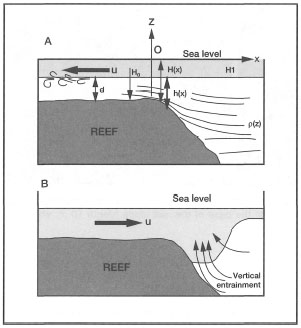
Figure 10.17. Possible model of localized upwelling induced by high-velocity tidal currents (tidally-induced upwelling on Buliulin Reef). A) Upwelling by tidal suction during flood tide. B) Vertical entrainment by tidal jet during ebb flow.
After Wolanski 1992.
However, the flourishing coral communities on both sides of the reef do not extend below the 10 m isobath, where there is an abrupt transition from hard substrate to fine carbonate sediments mixed with fluvial deposits. Thus, the depth extent of the coral communities seems to be substrate-limited rather than water quality or light-related. Based on 10-minute manta-tow surveys around the periphery of the reef, it became apparent that the southeast sector of the reef had a significantly patchier distribution of coral communities, when compared to the north, east and west sectors. The patchiness at the southeast sector of the Buliulin Reef was attributed to numerous shallow channels, which were the main outlets for the reef flat water during the ebb flow (fig. 10.18).
The location of the channels corresponds well with the predominant net southward flow in the region. Based on these limited observations, we may nonetheless speculate that low salinity is not the main limiting factor in this region of the archipelago. Turbidity and sedimentation associated with river runoff, or with highly productive reef flats, are most probably the dominant environmental factors determining the regional and local distribution of coral communities and coral reefs.

Figure 10.18. A) Sediment channel at the southeast sector of Buliulin Reef at high tide (depth 3 m). B). Sediment accumulation occurs at the base of the reef slope (depth 10 m; ebb flow). Note the greatly reduced water clarity.
Effects of High Salinities
Sea surface salinities are also influenced by evaporation which is controlled by solar radiation, by the regional and local wind fields, and by humidity. During the dry season, weather conditions may result in rapid heating and evaporation of extensive shallow intertidal pools and shallow lagoons, thus substantially increasing sea surface salinities. Under these conditions the formation of high-temperature (>31°C) and-salinity (>36 psu) water in the shallow reef flat pools may have a significant influence on the lower reef slope coral communities through a cascading mechanism (i.e., high-density water will spill over the reef slope). Downwelling of warm, saline, sediment-laden, reef flat water masses is a common occurrence, especially on reefs with extensive reef flats and steep drop-offs. For example, during a survey of an offshore reef on the northeast coast of Koon Island (southeast Seram), strong local downwelling was documented along the reef wall at a depth of 10 m. The circulation in this area is dominated by strong tidal currents that can reach velocities of up to 3 m.sec-1. The downwelling was characterized by a sharp change in temperature, from 28.2°C to 30.1°C, and a strong downward current with a velocity of about 0.5 m.sec-1. The turbid water was restricted to within 20 m of the reef wall, and provided a sharp contrast to the off-reef oceanic waters. The downwelling cell extended for about 300 m along the reef wall, where it stopped as abruptly as it had begun. Large feeding aggregations of planktivorous fish (e.g., Chromis amboinensis, C. atripes, C. xanthochir, C. analis, Pseudanthias luzonensis, P. dispar, P. lori) were oriented in an up-reef position, against the downwelling, in sharp contrast to the horizontal orientation of large schools of Pterocaesio randalli feeding about 15 m off the reef wall.
During intense ENSO events, these normal downwelling episodes may be greatly intensified in certain regions (e.g., Java Sea), triggering reef-bleaching events, which may be followed by substantial coral mortality (Brown and Suharsono 1990). Furthermore, the downwelling may be an important mechanism for the transport of fine-grained, shallow-water reefal sediments into deeper parts of the reef slope. Sediment cascading down the reef slope was observed on numerous occasions on most oceanic platform reefs in the Banda Sea.
LIGHT AND CORAL REEFS
The ability of corals to build reefs using the energy of the sun, is the key to the existence of all modern coral reefs, and perhaps all reefs in all geological time—VERON 1995
The statement certainly rings true, considering the fact that the coral-dinoflagellate holobiont is an autotrophic biological unit, whose complex biochemical processes are clearly light-dependent (Lewis 1974c; Jaubert 1977; Gattuso and Jaubert 1984; Kinzie 1987; Titlyanov 1987; Waymann et al. 1987; Falkowski et al. 1990; Harland et al. 1992). Perhaps with the exception of some temperate species (e.g., Astrangia danae), coral-zooxanthellae symbiosis is an obligate association, since all zooxanthellae corals rely to a great extent on their algal symbionts for nutrition (i.e., reef-building corals can derive up to 95% of all nutritional requirements from the zooxanthellae), and, most importantly, calcification (Porter 1976; Crossland et al. 1980b; Muscatine et al. 1981; Davies 1984; Muscatine 1990). It is, therefore, not surprising that solar energy is one of the key environmental factors limiting the distribution of zooxanthellate corals, and coral reefs in general.
Sunlight is the primary force that drives coral reef ecosystems, as well as all other autotrophic systems on the planet. In fact, with the exception of the chemosynthetic (i.e., chemical energy) deep oceanic benthic communities associated with hydro thermal vents, the earth's biosphere is driven by the Sun's energy. In simple terms, the basic fuel for the coral-zooxanthellae holobiont is the Sun's electromagnetic radiation which can be used by the zooxanthellae. The basic aspects of light that will have an effect on the coral symbionts is light quantity (the amount of energy delivered) and light quality (spectral composition), both of which are affected by a variety of factors. Both quantity and quality of light are greatly influenced by seasonal changes in day length (i.e., amount of energy received) and the angle of incident sunlight (i.e., quality of energy received). Therefore both the amount and spectral quality of insolation will decrease with increasing latitude (fig. 10.19).
However, after reviewing the available literature, it is apparent that the amount of solar radiation reaching the coral communities at their northern and southern limits of distribution is clearly not a limiting factor, since corals have been observed to grow at depths of 30-40 m at latitudes 31° S and 35° N in clear-water environments (Veron 1995). When we look at the distribution of the total amount of solar energy being received at various regions on Earth, it becomes apparent that the expected latitudinal gradient is not as clearly defined as one would expect (fig. 10.20).
There are obvious differences between the Arctic and equatorial regions, but at lower latitudes (50° N - 50° S) the pattern is not clearly apparent. For example, Indonesia, straddling the equator, is expected to receive the highest doses of solar energy, but in fact receives less over the year than the Red Sea, which is situated at the northern limit of coral reef distribution in the Indian Ocean. The average yearly amount of solar energy that is being received in Indonesia, ranges from 155 to 230 W.m-2, well within the range being received by the coral communities situated at the southern limit of their Pacific distribution at Lord Howe Island (31° S). Yet these systems differ greatly in terms of reef development and coral species richness.
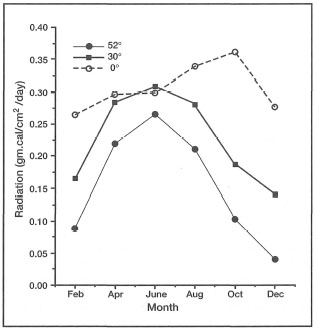
Figure 10.19. Average amounts of incident solar radiation over one year period, expressed as gm.cal.cm-2day-1 at latitudes 0°, 30° N and 52° N.
Source: Raymond 1963.
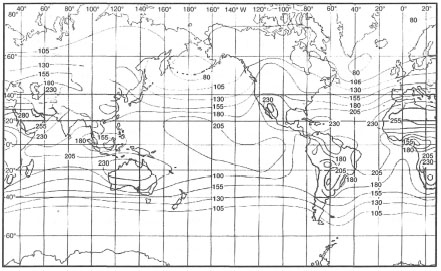
Figure 10.20. Average amount of solar radiation (W.m2) that is received at the earth's surface.
From Open University 1991b. Open. Univ. Ocean. Circ; p. 159; fig. 6.1.
The differences in the amount of solar energy being received at any one place on the planet, are closely linked with atmospheric conditions, which have a significant effect on insolation (i.e., the actual amount of solar energy reaching Earth's surface). As a general rule, large continental areas, on average, receive more insolation than oceanic regions, especially in the equatorial belt. The major factor responsible for lower insolation over equatorial oceans, as well as over tropical rain forests, is rapid evaporation which results in cloud formation. A higher concentration of water vapour in the atmosphere, and formation of clouds over most equatorial regions significantly increase light scattering, absorption and reflection, which ultimately results in reduced insolation. These processes are fundamental to global, regional and local energy distribution patterns along both spatial and temporal scales.
On any given clear sunny day in the tropical latitudes, the sea surface around noon receives, on average, 2500 μE.m-2.sec-1 of photosynthetically active (available) radiation (PAR), which can potentially be utilized in algal photosynthesis. However, not all of this surface energy is available to marine autotrophs, since the moment light reaches the sea surface it is quantitatively and qualitatively altered by numerous physical processes. Only a small fraction of the available solar energy reaches the autotrophs, and an even smaller fraction is actually used in photosynthesis.
The absorbed fraction is usually referred to as the photosynthetically usable radiation (PUR), and for zooxanthellae, and other marine algae, it is restricted mainly to radiant energy between 400 to 700 nm (i.e., nm = 10-9 m). The amount of PUR actually used in photosynthesis varies among marine autotrophs, and depends entirely on the composition of their photosynthetic pigments (e.g., chlorophyll), which determines the wavelength-specific absorption spectrum of the photosynthetic cells (Falkowski et al. 1990). Zooxanthellae have two absorption peaks within the PAR range. A wide absorption peak occurs between 400 and 550 nm, while a second narrower peak occurs between 650 and 700 nm.
While solar radiation does not seem to be a limiting factor in the latitudinal distribution of coral reefs, it is the key factor that determines their depth distribution (Bak and Luckhurst 1980; Sheppard 1982; Huston 1985). Considering the fact that the majority of zooxanthellate corals are restricted to the euphotic zone, it may be said that the key limiting factor with regards to depth distribution is symbiosis itself. Even under optimum conditions (i.e., clear oceanic water), zooxanthellate corals are generally restricted to depths between the surface and 100 m (Wells 1957, 1969), however, the most vigorous coral reef development seems to occur at depths of 3 to 20 m. In most tropical seas (e.g., Flores Sea), the 18°C isotherm lies roughly at depths between 150-200 m, which is below the euphotic zone even in the clearest ocean. Temperature is therefore not a major factor limiting depth distribution of corals in most tropical seas; however, it may be a factor in some upwelling regions (e.g., Galapagos).
On the other hand, symbiosis is most likely the key factor that has allowed the zooxanthellate corals to widely disperse throughout the oligotrophic tropical oceanic regions, although their dependence on symbiosis seems to severely limit their competitive ability, and possibly wider distribution, in high-latitude macroalgae-dominated environments (Crossland 1982,1988; Hatcher 1991,1993; Veron 1995). The role of symbiosis in environments with an abundant food supply (i.e., upwelling areas characterized by a high abundance of zooplankton) has not been investigated. The nature of the symbiotic relationship is discussed in chapter 7.
Light Environment in Indonesia
Coral reefs within the Indonesian Archipelago are situated in an equatorial region, and therefore, the question of whether light has an effect on the horizontal distribution patterns within the archipelagic seas may seem to be a non-issue. However, as was discussed earlier, the archipelago is not climatically homogeneous. In fact, there are significant climatic differences between the western (i.e., Sumatra, Java, Kalimantan and most of North and Central Sulawesi), and eastern (i.e., east of Java, except Irian Jaya) regions of the archipelago. One of the most obvious differences is the average annual rainfall, which is loosely indicative of the amount of insolation. The southeastern regions, on average, receive considerably higher insolation (between 200-230 W.m-2) than the western regions (150-190 W.m-2). These differences may be related to the drier, at times arid, climate of the Lesser Sunda Islands and southern Moluccas, a wide region influenced by the Australian continental mass to the south. In contrast, the mountain systems and extensive rain forests of Kalimantan, Sumatra, and Malaysia have a combined regional influence on cloud formation in the area, thus the climate is slightly more humid and cooler than in the eastern regions. Whether there is an ecologically meaningful correlation between insolation and coral reef distribution patterns within the archipelago remains to be seen, but it is not likely.
The actual amount of insolation actually reaching the various seas of the archipelago is not the main issue, since as far as coral light requirements are concerned, all the regions receive an excess of solar energy year-round. What is, however, important, is how much of the available insolation actually reaches the autotrophic communities, whether they are benthic or planktonic. The amount and quality of available light that reaches the autotrophs in the sea, depends on the synergistic effects of a number of environmental factors, and not just on the amount of insolation. Much of the incident light at the sea surface is reflected back into the atmosphere as a result of surface irregularities caused by waves (i.e., a function of the wind) and the angle of the sun. Even in the clearest of ocean water, 65% of irradiance that actually penetrates the sea surface is lost within the first metre of the water column, with only 1% reaching 60 m, and at about 200 m, less than 0.1% remains. Under ideal oceanic conditions, light penetration is primarily controlled by the optical properties of seawater, and by water column productivity. This certainly is the case in highly productive oceanic systems, such as the Banda Sea, where light penetration (i.e., as measured by a Secchi disk) has a distinct seasonal pattern that is primarily related to seasonal fluctuations in primary and secondary production rates (fig. 10.21). The attenuation of light (through absorption and scattering) as it passes through the water column in coastal waters is influenced by a variety of other factors that will be discussed in the following section.
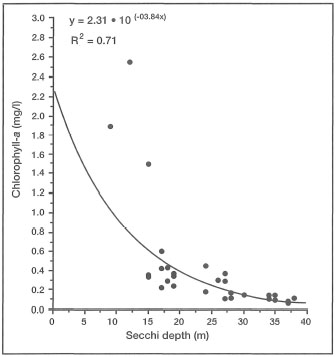
Figure 10.21. Correlation between water transparency as measured by Secchi disc (m), and primary production as measured by chlorophyll-a concentrations, Banda Sea.
Source: Gieskes et al. 1989.
While the differences in insolation among the various regions of the archipelago are relatively minor, and most likely biologically and ecologically insignificant, as far as reef and coral distributions are concerned, the amount of solar energy reaching benthic communities clearly varies among regions. Higher rainfall and different geological setting in the western regions of the archipelago, and Irian Jaya, are two key factors (we shall ignore anthropogenic factors for now) that may have a significant influence on both the quantity and quality of land runoff that enters the coastal waters. In terms of coral distribution patterns, land runoff is of primary importance, since the suspended particulate load which it carries, directly influences light penetration and ultimately the amount of light reaching the benthic autotrophic communities.
Turbidity and Suspended Particulate Matter.
DEFINITIONS. Turbidity can simply be defined as water clarity or transparency. However, while being governed by the optical properties of seawater, turbidity is mainly a function of suspended particulate matter (SPM) concentrations and dissolved organic compounds in the water column. There is an obvious positive relationship between turbidity and SPM, but the synergistic effects are complicated. Turbidity mainly relates to light-dependent processes, while SPM, in addition to directly affecting turbidity, has a number of other effects that should be treated separately. Clear separation between the effects of turbidity and SPM are difficult, and rarely studied in corals (Yamazoto 1986).
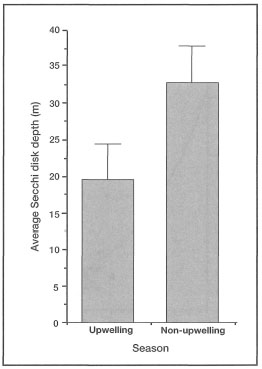
Figure 10.22. Comparison of water clarity (means ± STD) between the Northwest Monsoon, a period of downwelling, and the Eastern Monsoon, a period of upwelling, in the Banda Sea. Lower water clarity is primarily a function of high phytoplankton and zooplankton biomass during upwelling (May-September). Conversely, high water transparency is directly related to low primary productivity during the Northwest Monsoon (November-April).
Source: Gieskes et al. 1989.
Suspended particulate matter can be either biotic (e.g., phytoplankton, zooplankton, organic detritus, etc.) or inorganic (e.g., silt, sand, etc.) in composition. In oceanic environments, removed from the influence of continental processes, turbidity is generally low (i.e., low SPM concentrations; <0.5 mg/l). This may be true for the vast regions of the central Pacific Ocean, but the oceanic environments within the Indonesian Archipelago (e.g., Banda Sea) experience marked seasonal fluctuations in turbidity (i.e., SPM) (fig. 10.22). These unusual conditions are primarily a result of seasonal plankton blooms and overall increases in primary production associated with pulses of upwelled nutrients during the Southeast Monsoon (Gieskes et al. 1989).
The moment light penetrates the sea surface, its intensity starts to decrease exponentially with increasing depth as a result of absorption and scattering by water molecules. However, seawater also contains various biotic and abiotic compounds, as well as a multitude of living organisms, that significantly alter the absorption and scattering properties of water masses. The net result of all the particles in the seawater is lower water clarity, or higher turbidity, when compared to pure water. As more particles are added to seawater, turbidity continues to increase and less light reaches both planktonic and benthic communities (fig. 10.23).
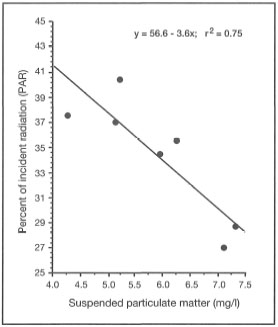
Figure 10.23. Regression of percentage of incident light (photosynthetically available radiation - PAR) on total suspended particulate matter concentrations (mg/l).
Source: Tomascik and Sander 1985.
The absorption and scattering of light in turbid coastal waters has a greater effect on the depth of the euphotic zone, than in open ocean conditions, since higher turbidity not only reduces the amount of solar radiation, but also causes a major shift in spectral quality of light reaching the benthic communities (fig. 10.24). The differences in spectral quality between oceanic and coastal environments are related to the composition of the SPM and dissolved organic substances present in the seawater. Unlike the oceanic waters, where the maximum energy penetration (up to 200 m) is in the blue part (480 nm) of the light spectra (i.e., oceanic water looks blue), the maximum penetration in turbid coastal waters is shifted to the yellow (550 nm), with significantly lower penetration depth.
SOURCES AND COMPOSITION OF SPM.
Oceanic Environments. In oceanic regions suspended particulate matter is composed almost entirely of organic matter, which consists mainly of bacterioplankton, phytoplankton, zooplankton, various metabolic by-products (e.g., larvacean mucus 454 sheets, calcareous and siliceous tests, etc.), and organic detritus. These biotic and abiotic components usually form large aggregates (>0.5 mm in diameter) in the water column, and are commonly referred to as marine snow (Shanks and Trent 1979; Alldredge and Silver 1988). Associated with SPM and marine snow are various dissolved organic substances which are either metabolic wastes (e.g., fecal pellets), or products of organic decomposition. The synergistic effect of SPM and dissolved substances in the water column plays a significant role in light extinction.
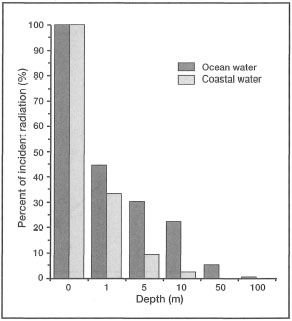
Figure 10.24. Percentage of total incident solar radiation at various depths in clear oceanic water and turbid coastal water.
Source: Raymond 1963.
While high concentrations of SPM, and therefore turbidity, may have negative impacts on autotrophs, the nutrient-rich marine snow (comprising most of SPM) is an important source of food for pelagic fauna (e.g., zooplankton and ichthyoplankton) (Shanks and Trent 1979; Alldredge and Silver 1988). As oceanic water passes over the reef, SPM concentrations increase as a direct result of increased productivity (benthic and pelagic), particularly in lagoonal waters where relatively turbid conditions predominate.
Oceanic and reefal detrital SPM is an important source of food for benthic filter and suspension-(particle) feeding organisms, and perhaps planktivorous reef fish as well. Furthermore, much of this drifting organic matter can be utilized directly by corals themselves (Lewis and Price 1975; Sorokin 1978). While being an autotroph [photosynthesis-to-respiration ratio (P/R) >1], the coral-zooxanthellae holobiont is also a highly efficient predator, equipped with a variety of auxiliary-feeding habits (Muscatine and Porter 1977), one of which is particulate or suspension-feeding (Sorokin 1973; Lewis and Price 1975; Lewis 1977b).
Earlier studies in the Banda Sea (Nontji 1975) indicated that seasonal fluctuations in primary production, and therefore water clarity, were monsoon-driven and associated with upwelling (Wyrtki 1961). Lowest transparency occurs during the Southeast Monsoon (May-September), when strong easterly winds set up a large-scale upwelling system that significantly increases phytoplankton and zooplankton biomass in the water column. During the Northwest Monsoon, which is a period of general downwelling (i.e., sinking) (Wyrtki 1961), the water clarity improves considerably as both phytoplankton and zooplankton biomass drop.
At the peak of upwelling, high primary and secondary production in the water column considerably reduces water clarity and thus the depth of the euphotic zone. While higher productivity may be beneficial for coral communities in general, especially at depths of less than 20 m, significant reduction in water transparency during the upwelling period may effectively reduce the maximum depth limit of reef-building. In the productive coastal waters, light penetration becomes a significant factor in both the horizontal (regional to local) and vertical distribution of coral communities.
Coastal Environments. In comparison, most coastal waters in the archipelago are characterized by high turbidity, and the suspended particulate matter is of various biotic and abiotic origins, both aquatic and terrestrial. However, in addition to the SPM, coastal waters are known to contain various soluble pigments or 'yellow substances' that are responsible for significant absorption of light (Jerlov 1951).
In tropical coastal environments, the bulk of the dissolved organic acids and other substances originate from mangrove forests and tidal swamplands. The influence of terrestrial runoff on inshore coral reef communities has been clearly demonstrated in some areas, by the detection of humic and fluvic acids in the skeletons of massive species of Pontes (Isdale 1984; Boto and Isdale 1985; Susie et al. 1991). On the Great Barrier Reef, humic acids have been detected as far as 80 km offshore, with highest concentrations being recorded in the inshore coastal waters. The main source of humic and fluvic acids is terrestrial plant matter, and according to Susie et al. (1991), soils of tropical rain forests are particularly rich in these organic compounds. Humic acids are continually being leached into the soil and to small, streams that eventually discharge into the coastal waters. However, during heavy rainfall, tropical rain forests lose a considerable amount of surface soil material (contrary to popular belief) that may be particularly rich in these substances. The work by P. Isdale and his colleagues at the Australian Institute of Marine Science (AIMS) clearly demonstrated that corals (i.e., Pontes spp.) are able to incorporate humic acids into their skeletons during calcification.
Since humic acid concentrations in most tropical coastal waters are related to the annual patterns of rainfall (i.e., wet-dry), and river runoff, the skeletons of massive corals-when exposed to UV light-exhibit distinct yellow-green fluorescent bands. The periodicity of the fluorescent bands is closely correlated with the amount and frequency of rainfall and the intensity (i.e., volume) of river discharge (Susie et al. 1991). Humic acid concentrations in the coastal waters are, on average, lower during the dry season than during the wet season. During the wet season, high concentrations frequently occur following periods of heavy rainfall. This discovery has helped the Australian researchers to reconstruct past river discharge volumes, and to discern patterns of anthropogenic influences in the main drainage areas bordering the Great Barrier Reef.
In contrast, comparable success with fluorescent banding in Indonesia has not materialized (Scoffin 1986). However, it should be noted that fluorescent banding studies have been restricted mainly to coral reef areas along the north coast of Java (i.e.,Kepulauan Seribu and Jepara). In all studies conducted so far, clear bright UV banding is visible in all coral skeletons, but seasonal patterns are not discernible (Scoffin 1986). UV-banding patterns in corals from Kepulauan Seribu surprisingly show no significant correlation with mainland rainfall data, as they do on the Great Barrier Reef. At first this may be surprising, considering the fact that most parts of the archipelago, including Java, are under the influence of the monsoonal climate with distinct dry and wet seasons. Seasonal periodicity in the volume of river discharge has been documented for some rivers (e.g., Solo and Brantas Rivers in East Java), and therefore, one would expect to find strong fluorescent banding patterns in most inshore corals. Closer examination reveals, however, that the main study sites are located in coastal areas severely impacted upon by anthropogenic influences. Of significance are the greatly altered coastal drainage patterns and the deforestation (agricultural conversion) of the tropical forests in source watersheds. Seasonal UV banding in corals along the north coast of Java, which may well have existed under pristine environmental conditions in the distant past, is confounded by anthropogenic influences that have greatly modified previous natural environmental conditions along much of the coastline. If UV fluorescent banding in massive corals is indeed closely linked to land runoff, as seems to be the case in Australia, then the lack of banding in Kepulauan Seribu corals may indicate major changes in coastal environment have occurred, and corals are now subjected to the effects of continental processes at higher intensities and frequencies.
Effects on Coral Reef Distribution. It has often been stated, and is a widely held view in popular literature, that coral reefs have evolved in clear and oligotrophic (i.e., low nutrient availability) oceanic waters. An often-stated analogy is that a coral reef living in clear nutrient-poor tropical oceanic waters is like an oasis in the desert. This maybe true, but only for the 'blue-water' oceanic reefs (i.e., atolls) of the Pacific and Indian Oceans. Unfortunately, the direct corollary of this assertion is that vigorous coral reef development occurs mainly under the above 'presumed' ideal oceanic conditions, which implies that coral reefs in the more turbid coastal regions are living under less-than-optimal conditions. From the resource management and conservation perspective, this is a potentially dangerous view, since, at least from the Indonesian experience, these turbid, yet highly productive, areas are often ignored and excluded from management initiatives. Indeed, conservation efforts in Indonesia are too often, if not exclusively, directed at the traditionally 'blue-water reefs' (i.e., high tourism potential) of the eastern regions.
As was clearly pointed out by Potts (1983), about 50% of all Indo-Pacific reefs are found along shallow continental shelf margins. An unknown percentage of these reefs are scattered throughout the shallow continental seas of the Indonesian Archipelago (e.g., Java Sea, Natuna Sea, Arafura Sea), and along still relatively undocumented coastlines of Kalimantan, Irian Jaya and other large islands. Water clarity along the coastlines of these large islands is considerably less when compared to the oceanic waters of the Banda and Flores Seas, where light penetration is high. For example, the extinction coefficient (k') along the northwest coast of Java ranges between 0.85 m-1 and 1.7 m-1, while in the west Java Sea k' ranges between 0.34 m-1 to 0.09 m-1. In comparison, k' in the Banda Sea varies between 0.09 m-1 during the upwelling season, and 0.05 m-1 during the Northwest Monsoon when upwelling is inhibited by predominantly westerly winds. It is unfortunate, that, by focusing most research on the 'classical' blue-water reefs of the Indo-Pacific (i.e., atolls), a few generalizations (e.g., low nutrients, high water clarity) have permeated the popular literature, even though widely read papers have provided solid evidence that coral reefs have a remarkable capacity to develop and 'flourish' under relatively turbid conditions (Verwey 1931a).
But we see that even in the tropics, so close to the equator, and even in a coral reef region, we need not always have the blue clear water one dreams of in thinking about reef formation.—VERWEY 1931A
Turbidity is, however, considered to be more important than salinity in inhibiting reef development, even in regions where other environmental factors, such as temperature and substrate, are favourable (Guilcher 1988). As was pointed out earlier, the coral-reef-depauperate coastlines of Southeast Asia, India and the northwest coast of Brazil are influenced by turbidity plumes from major river systems, which confound or override the effects of salinity. In Indonesia, there is a distinct longitudinal gradient of water clarity which may have some influence on reef distribution and development (fig. 10.25).
Comparative coral reef studies along this longitudinal gradient have not been conducted, and the results of individual reef surveys in the various regions are not comparable because of varying methodologies and reporting. While some coral data are now available, a common deficiency is the lack of an environmental component. Thus, while we may discern significant differences between different regions in terms of coral community structure (i.e., diversity or coral cover), the main question as to why these differences occur remains unanswered. Nonetheless, the effects of turbidity on the coral community structure and function are well-known and have been documented elsewhere (fig. 10.26).
In many coastal areas of the Indonesian Archipelago coral reefs flourish or persist, albeit within a very narrow depth range, under turbid conditions (e.g., k'=1.0). As an extreme example let's examine the coral reefs of Pulau Panjang which are located in Banten Bay (West Java). Coastal waters in Banten Bay are muddy brown most of the time, with conditions improving as one moves further offshore. Turbidity in the bay is high and a Secchi disk disappears in less than 2 m. The entire Banten Bay is influenced by heavy land runoff from intensively used agricultural lands, which has considerably increased soil erosion, and therefore, sedimentation rates in the bay area (Kiswara 1992). However, all islands in the bay are fringed by reefs. The intertidal reef flats are dominated by seagrass communities consisting mainly of Cymodocea serrulata, Halophila ovalis, H. uninervis, and Thalassia hemprichii, which, however, are under considerable siltation stress (Kiswara 1992). The seaward edge of the reef flat (1.5-2 m below sea level) is the only zone on the reef where corals are able to survive without visible trauma, and where visible recruitment was observed (i.e., abundance of small, <5 cm colonies). However, major reef-builders such as Acropora are almost absent, and only a few colonies of Acropora millepora, A. aspera, and A. nobilis remain. The community consists of encrusting forms dominated by small colonies of Pontes spp., Goniopora spp., Pavona varians, and Leptoseris spp. as well as Fungia (Verrillofungia) concinna, F. (Verrillofungia) repanda and (Fungia) fungites. Down the reef slope, the live coral zone extends to a depth of only 2.5 m, where a sharp transition occurs from a solid carbonate substrate to fine silt deposits that are heavily burrowed by macroinvertebrates. At the transition are a few monospecific stands of Acropora nobilis. The reef slope community consists of Montipora spp., Favia spp., Favites spp., Galaxea spp., Pectinia spp., Oulastrea spp., Pachyseris spp. and Merulina spp. Most corals show signs of 'stress' (i.e., white spots, black spots, copious mucus production, and tissue sloughing). Horizontal visibility at 1 m depth is less than 3 m. Bioturbation is a significant source of fine resuspended sediments, which are a major contributor to the reduced water clarity. The narrow reef slope is dominated by Turbinaria spp., Montipora spp., Pocillopora spp., Galaxea spp., Pontes spp. and Goniastrea spp. The origin of the fine reef slope sediment is not certain, but it may originate from the reef flat. The sediments at the base of the reef slope (5-8 m) consist of fine sand and mud. The muddy sediments support a relatively diverse assemblage of soft-bottom benthic fauna consisting of about 121 species, with polychaetes being the dominant taxonomic group (i.e., 69 species) (Kastoro 1992).
Figure 10.25. Comparison of average yearly (± STD) Secchi disc readings (m) along a longitudinal gradient from Sunda Strait to Arafura Sea.
Figure 10.26. Regression of coral diversity (Brillouin's diversity index H) on total suspended particulate matter (mg/l).
Source: Tomascik and Sander 1985, 1987.
Effects of SPM. In addition to affecting light-dependent metabolic processes (i.e., photosynthesis) through turbidity, SPM has a much wider range of effects that influence the functional integrity of coral communities. While the effects of sedimentation on corals and coral reefs have been studied, and reviewed extensively (Marshal and Orr 1931; Hubbard and Pocock 1972; Dodge and Vaisnys 1977; Wijsman-Best et al. 1981; Brown and Howard 1985; Hodgson 1988; Stafford-Smith 1990, 1993; van Woesik 1994), the effects of SPM have been largely neglected (Yamazoto 1986), even though a few studies have demonstrated that coral community structure (Cortes and Risk 1985; Tomascik and Sander 1987b) arid coral growth rates (Tomascik and Sander 1985) are highly susceptible to high SPM concentrations (fig. 10.27).
Chansang et al. (1993) have recently shown that the growth rates of Pontes spp. during the rainy season (i.e., maximum SPM and turbidity) were 0.62 mm per month, and 1.54 mm per month during the dry season when SPM and turbidity are lower. Similar response was recorded in Acropora formosa, with growth rates ranging between 3.0 to 5.7 mm per month during the rainy and dry seasons, respectively. Similar conclusions can be drawn from laboratory studies, where, for example, Yamazoto (1986) demonstrated that the survival rate in corals is greatly reduced by an increased load of SPM in the water column (fig. 10.28). Yamazoto (1986) attributed the reduced survival time under high SPM concentrations to four possible effects: 1) direct abrasion through physical contact; 2) suffocation (direct or indirect); 3) energy expenditure in production of mucus and ciliary movement; and 4) starvation due to food dilution. Which of these effects is dominant, will depend entirely on the nature and quality of the SPM and the local current regime (Verwey 1931a). High concentrations of relatively soft, organically-rich detritus, that is often associated with sewage discharge, will have a different effect on coral and coral communities than, for example, high concentrations of SPM consisting mainly of large reefal or non-reefal inorganic sediments. In shallow, tropical coastal waters, organic compounds makeup a substantial bulk of suspended particulate matter. In the relatively oligotrophic waters of the eastern Caribbean, for example, organic matter constitutes 30%-50% of suspended particulate matter, with higher values recorded in areas heavily influenced by anthropogenic activities (e.g., tourism, sewage discharge, land runoff, etc.) (Tomascik and Sander 1985). The organic fraction of SPM may represent an important pool of nutrients (Lewis 1977b), and up to certain concentrations, organically-rich SPM may promote coral growth (Tomascik and Sander 1985).
Figure 10.27. Regression of coral growth rates (cm/yr) on total SPM (mg/l).
Source: Tomascik and Sander 1985.
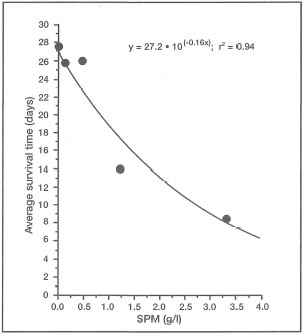
Figure 10.28. Regression of survival time of Pocillopora on SPM (mg/l).
Source: Yamazoto 1986.
In fact, organically-rich SPM in the water column may be viewed as a distinct ecosystem or microcosm, since each individual particle, however small, is associated with bacterial fauna, and frequently with phytoplankton and zooplankton as well (Alldredge and Cohen 1987). Most of the SPM in coastal waters, especially in coral reef areas, consists of various aggregates that are clearly visible. The nature of the organic detritus is varied, but the main components are various bits-and-pieces of benthic algae, seagrasses, periphyton and flocculated coral mucus that is a prime media for bacteria (Marshal 1968; Gerber and Marshal 1974; Marshall et al. 1975; Westrum and Meyers 1978; Ducklow and Mitchell 1979; Lewis 1979; Gottfried and Roman 1983; Coffroth 1984). Heterotrophic marine bacteria are responsible for rapid decomposition of most organic particulate matter in tropical climates, and therefore their metabolic activity has a direct effect on biological oxygen demand, which may lead to a reduction of dissolved oxygen concentrations in the water column during early morning hours. Figure 10.29 demonstrates the significant positive relationship between organic matter concentrations in the water column and biological oxygen demand.
At what concentrations SPM becomes harmful to corals and coral communities, and thus detrimental to reef development, will depend on the nature of the SPM, the type of reef community and the local environmental conditions (i.e., currents, temperature, salinity, oxygen, etc.). While these are important issues for future coral reef management policies, very little research is being conducted to answer these questions. Whether or not a sewage plant will have a greater impact on the coral community than, for example, discharge of raw sewage, needs to be addressed on a case-by-case basis. It is likely that many coral communities along the turbid coastal waters in the western part of the archipelago (e.g., Banten Bay, Jepara, Jakarta Bay) are living at the upper limits of tolerance with regards to turbidity levels and SPM concentrations. In fact, Jakarta Bay is a good example of an area where turbidity played a key role in the demise of most inshore reefs.
In Jakarta Bay, SPM concentrations measured during both the Northwest (i.e., wet) and Southeast (i.e., dry) Monsoons, range from 5-45 mg/l, depending on the distance from shore (Nontji and Supangat 1977). Highest SPM concentrations in Jakarta Bay occur during the Northwest Monsoon (December to February), the period with the highest rainfall in West Java. However, in the offshore waters (3-6 km), seasonal differences in SPM concentrations are not statistically discernible. In fact, SPM concentrations, and thus turbidity, may increase during the Southeast Monsoon (i.e., dry season), especially in the eastern regions of the Java Sea. One possible explanation is that during the Southeast Monsoon, surface currents in the Java, Flores and Banda Seas are flowing in a westward direction driven by strong easterly winds. These carry with them high phytoplankton and zooplankton biomass, which originate from the upwelling regions of the Banda and Flores Seas (Nontji and Supangat 1977; Nontji 1978; Setiapermana et al. 1980). The lack of seasonality in SPM concentrations in the Java Sea may be related to this transport. Unfortunately, the database is not extensive enough to verify this.
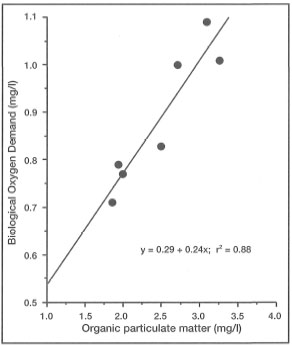
Figure 10.29. Regression of biological oxygen demand (BOD mg/l) and suspended organic particulate matter (mg/l).
Source: Tomascik and Sander 1985.
High SPM concentrations in the coastal waters during the Northwest Monsoon seem to indicate that a close relationship between SPM and land runoff should predominate, and that most of the SPM during this period should be of terrestrial origin. The terrestrial, or river-borne, origin of suspended particulate matter in Jakarta Bay during the Northwest Monsoon is implicated by a strong inverse relationship between SPM concentrations and sea surface salinities (fig. 10.30) (Nontji and Supangat 1977). The offshore SPM gradients that exists during the Northwest and Southeast Monsoons are clearly demonstrated in figure 10.31. All existing coral communities, and what remains of the reefs in Jakarta Bay (i.e., Nyamuk Besar and Kecil, Damar Besar and Kecil, Cipir, Kelor, Air Besar and Kecil, Ubi Besar, Untung Jawa, Rambut, Dapur, Bokor, and Bidadari) are thus influenced by high turbidity. Survival of many of these coral communities is linked to the strong currents that prevent sedimentation of the fine suspended particulate matter. The important role of currents in maintaining flourishing coral reef communities in the turbid Jakarta Bay has been recognized in the past (Verwey 1931a). Working on Onrus Island (Pulau Eapal), Verwey (1931a) came to the conclusion that turbidity, provided strong currents are available, will have mainly a limiting effect on the depth distribution of the live reef, whereas, without the currents, sedimentation will smother and kill the reef.
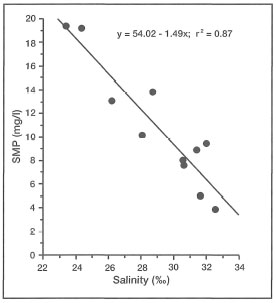
Figure 10.30. Regression of SPM (mg/l) on salinity (ppt) in Jakarta Bay. The strong negative relationship suggests terrestrial origin of the SPM during the wet season.
Source: Nontji and Supangat 1977.
Turbidity Induced by Tidal Upwelling. Koon Case Study. In most offshore regions, especially where extensive reef development occurs, seawater turbidity is usually at the lowest, and therefore an often neglected parameter measured during ecological studies. The following example illustrates why the low-turbidity assumption for oceanic reefs may not be as widely applicable for the offshore reefs in the Indonesian Archipelago.
An extensive reef complex begins at the southeastern tip of Seram, and extends in an easterly direction for 60 km into the northeastern Banda Sea (fig. 10.32).
The reef complex is situated on a submarine ridge that rises from a depth of about 500 m, and can be viewed as an extension of Seram. The submarine ridge is part of the non-volcanic Outer Banda Arc that skirts the Weber Deep along its eastern margin (van Bemmelen 1949). The complex consists of three main reefs that are separated by relatively shallow (maximum depth 30 m) 1-km-wide channels. The reef flat has a maximum width of 6 km, with two large islands on its western margin. Seramlaut and Geser consist of calcareous sandstone of Mesozoic origin and raised coral reefs (van Bemmelen 1949). The reef complex forms an impressive natural barrier between the Seram and Banda Seas, located to the north and south of the barrier, respectively. The easternmost extent of the reef is about 60 km from Seram, and therefore, sufficiently removed from any terrestrial influence. The reef flat in this region is about 2 km wide. The area is influenced by strong tidal currents, which on a number of occasions were estimated at 2-3 m.sec-1. From the seaward edge of the reef flat (fig. 10.33), the reef slopes down to a depth of 5 m where it ends in a vertical drop-off.
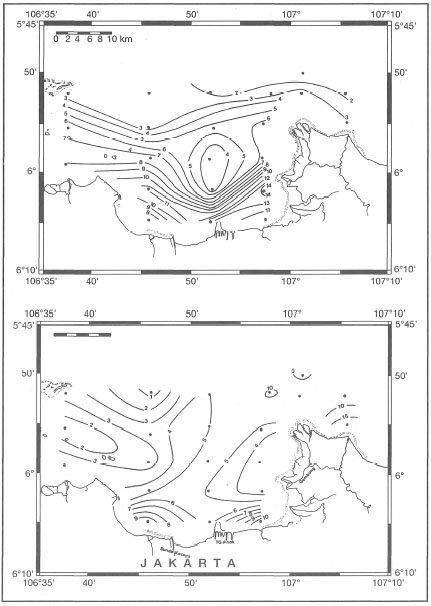
Figure 10.31. Horizontal SPM gradients in Jakarta Bay, during the Northwest (January) and Southeast (August) Monsoons. Values in mg.l-1.
From Setiapermana et al. 1980.
Figure 10.32. Hydrographic chart and location of Koon Island. Black arrow indicates location of dive site. Depth in metres. Kr - reef. (Chart No. 399).

Figure 10.33. The seaward margin (neap low tide) of an extensive intertidal reef flat with Koon Island, southeast Seram, in the background. Koon Island is a vegetated cay at a southern limit of a 60-km-long, barrier-like, reef complex up to 6 km wide.
Photo Tomas and Anmarie Tomascik.

Figure 10.34. Soft coral- and sponge-dominated coral reef community in a region where tidally-induced upwelling is an everyday phenomenon. The reef supports a high diversity of reef- and reef-associated fish fauna. Feeding "frenzies" by large planktivores (e.g., Odonus niger, Hemitaurichthys polylepis, Heniochus acuminatus, Naso brevirostris, Naso hexacanthus, Naso vlamingii) seem to occur during upwelling events.
Photo by Tomas and Anmarie Tomascik.
Our first impression of the reef site was that the environmental conditions were characteristic of an offshore platform reef, which should be dominated by a coral community characteristic of high-energy oceanic reefs (i.e., mainly Acropom spp.). However, in the region where the Seram and Banda Seas meet, the reef community is dominated by a diverse assemblage of soft corals and sponges, usually characteristic of more turbid areas. Even though the coral community is subordinate in terms of abundance, it is nonetheless highly diverse, with 100-150 species. Interestingly, many coral species found on the seaward reef slope were characteristic of more turbid lagoonal environments, and the monospecific stands were unusual, and strangely out of place. The most abundant species were Goniopora lobata, G. columna, G. tenuides, Pontes cylindrica, Acropora nobilis, A. formosa, A. abrolhosensis, and A. yongei. However, there were also large monospecific stands of species characteristic of high-energy environments, such as Acropora palifera, A. cuneata, A. robusta and A. danai. The area is relatively sheltered, and wave height seldom reaches 1 m (according to local fishermen). Horizontal visibility underwater can exceed 40 m, suggesting that SPM concentrations are extremely low. In addition, the reef supports an "amazing" abundance of reef fish and large schools of pelagics (fig. 10.34).
The reason for this high productivity, the unusual coral community and the filter-feeding benthos became clear to us during one of the dives. The ambient temperature measured during one of the survey dives, at a depth of 10 m on the seaward reef slope, was 28.0°C. The current was running in an easterly direction parallel to the reef wall at a velocity of about 1 m.sec-1. Horizontal visibility was about 35 m and vertical visibility exceeded 50 m when the reef wall disappeared into the "deep blue". The first sign of something unusual was a distinct blackening of the deep reef slope whose outline slowly disappeared in a black layer that was slowly moving up the reef wall. Shortly afterwards, a cold water front spilled over the shallow reef slope, which caused the temperature to drop to 25.2°C, almost 3°C below ambient. The mixing of the cold and warm reefal water masses resulted in an intense shimmering effect that reduced horizontal visibility to less than 10 m. At about the same time the current slowed considerably and changed direction to southeast, at an angle up and over the reef slope. Within 20 minutes of sighting the black layer below, the water suddenly turned deep green and the visibility dropped to about 4 m. There was a sudden feeding frenzy of large schools of planktivores just off the reef. The temperature remained at 25.2°C until the end of the dive.
The above example serves to illustrate that tidally-induced upwelling events may have an important role in the structuring of coral communities in offshore areas where tidal action is a dominant factor. According to the hydrological records for this region, water masses with temperatures of 25°C (between September and October) at the end of the Southeast Monsoon are restricted to depths of 75 to 90 m (Institute of Marine Research 1974). This would suggest that the upwelled water masses originated at depths below 50 m, which agrees with our visual observations. The change in water colour observed over the reef was associated with the upwelled water mass, and is most likely due to high concentrations of SPM consisting of chlorophyll and other pigments associated with the Deep Chlorophyll Maximum (DCM) layer. The DCM layer during this period was located at depths between 60-80 m (Institute of Marine Research 1974; Gieskes et al. 1988).
In addition to phytoplankton, the upwelled water most likely contains high concentrations of SPM (e.g., marine snow), which tends to concentrate along density gradients. Furthermore, the DCM layer is also associated with high zooplankton biomass. Planktonic foraminifera are known to aggregate at the DCM layer. The tidal currents provide sufficient turbulence in the area to facilitate coagulation and flocculation processes necessary for production of aggregates (Alldredge and Gotschalk 1988). Regardless of the depth, the upwelled water mass elicited a feeding behavior from planktivorous reef fish, and many coral colonies had their tentacles fully expanded. Note that tentacle expansion in corals may also be a response to high current velocities. It seems that the tidally-induced upwelling may be an important source of food supply for the reef community, and the dominance of filter-feeding benthos in the area may be the consequence. Whether the rapid shifts in temperature have an impact is not known.
Tidally-induced upwelling events are not unique to this particular area of the archipelago, and have been observed to occur in areas with strong tidal regimes (e.g., Berau Barrier Reef in East Kalimantan; Banda Islands; Lucipara Islands, and Kepulauan Seribu). Some of these areas offer new opportunities to study reef development and reef community structure under upwelling conditions that differ greatly from our classical perceptions (Glynn 1976).
Turbidity has been shown to vary naturally across the archipelago with identifiable longitudinal gradients. As expected, high turbidity is usually found in most coastal waters, especially in close proximity to rivers. From the available information, SPM concentrations are influenced by a number of natural processes, however, land runoff from anthropogenically impacted coastal areas is increasingly becoming the key limiting factor for many inshore coral reef communities. SPM concentrations vary greatly, as does turbidity, across the archipelago depending on the habitat, ranging from <0.5 mg/l in the clear oceanic regions of the Banda and Flores Seas during the Northwest Monsoon, to 10,000 mg/l (fluid mud) in the deltaic environments (Hoekstra 1989). These pronounced differences in turbidity and SPM among various regions of the archipelago are most likely the primary forces determining the distribution of inshore reef communities; however, systematic studies at an appropriate taxonomic level have not been conducted.
SEDIMENTATION
Definitions
Sedimentation can be defined as the act or process of depositing sediment by mechanical means from a state of suspension in water, or as a process of forming and accumulating sediment in layers (Bates and Jackson 1980). Too often, however, the term is used loosely to describe general environmental conditions in areas where turbidity is high, even though no actual data are usually presented. This is based on the generalization that high turbidity (high load of SPM) is directly related to sedimentation. The sedimentation rate of suspended particulate matter depends firstly on the physical structure of the particles themselves (i.e., particle volume, surface area perpendicular to the direction of sinking, density, and porosity), secondly on the physical properties of seawater (i.e., density), and finally on the hydrology of the area (i.e., current velocities, shear stress, vertical location of SPM in the water column, settling velocities of particles, and turbulent mixing). Sedimentation rates are usually measured with the aid of sediment traps, which vary greatly in construction and dimensions from study to study, and as a result few sedimentation studies are quantitatively comparable. In addition, environmental conditions that affect many of the above factors vary greatly from place to place, and thus interpretation of sediment-trap data can be problematic at best. Furthermore, in high-energy environments, such as coral reefs, sediment traps measure only the downward flux of suspended particulate matter, and not the actual accumulation of sediments in a particular place over time (which is a combination of downward flux and resuspension).
The above discussion may seem to be academic; however, a clear understanding of what is and what is not sedimentation has serious implications in coral reef monitoring studies designed to obtain environmental information for the formulation of conservation and management policies. More important, however, is the use of sediment traps in environmental impact assessment (EIA) studies. A clear understanding of turbidity and sedimentation is critical for the interpretation of results and the subsequent formulation of mitigating measures. It is possible to obtain high sedimentation rates in pristine areas with high water clarity if the region is influenced by rough weather; however, what is being measured is resuspension of existing sediments, an impact of short duration and within the tolerance limits of the coral community. On the other hand, low sedimentation rates are probable in very turbid areas where coastal currents keep the SPM in suspension. Thus, while the corals are being influenced by low light levels and perhaps physical abrasion, the sedimentation data may well indicate that sedimentation rates are below the levels measured at various reference stations. Based on this information an erroneous conclusion could be reached that the pristine site is impacted while the impacted site is not. This situation is not theoretical, and has been reported elsewhere (Tomascik and Sander 1985). In this particular study, average sedimentation rates in an area influenced by discharge from a rum refinery were 19.98 mg.cm-2.day-1 while at a reference site 14 km from the impact area, sedimentation rates were 37.19 mg.cm-2.day-1. In comparison, SPM concentrations at the impacted site were 7.32 mg.l-1, while at the reference station SPM concentrations were 4.26 mg.l-1. Based on the sediment trap data alone, a serious error may have been made with regards to the actual conditions.
Sediment Transport, Origin and Effects
Of primary concern in the present context is the origin of terrigenous sediments that are formed by natural continental processes. However, the sediments along the Indonesian coastlines can be either terrigenous in origin or products of biogenic processes, or a mixture of both. The terrigenous sediments are derived predominantly from the weathering and erosion of hard volcanic and other indurated rocks as well as soft and unconsolidated sedimentary and alluvial deposits. The lithology of the watershed has an important role in the composition of the river load, and to a great extent determines the composition of coastal sediments.
Terrigenous sediments are more predominant in coastal areas with higher rainfall. In these regions (e.g., north coast of Java, South Kalimantan), terrigenous muds and sands (rich in clay minerals) form extensive coastal intertidal mud flats, which support diverse and productive soft-bottom benthic communities. In the drier and non-volcanic regions of the archipelago (e.g., Outer Banda Arc), shallow-water coastal sediments are predominately biogenic carbonates as well as chemically precipitated carbonates. Carbonate sediments form vast intertidal flats and generally replace the clay-rich muds in the estuaries. In other regions, such as young volcanic islands surrounded by fringing reefs, coastal sediments may consist of a mixture of volcanic material and carbonate sands. The role of waves and longshore currents in the distribution of sediments along shorelines, will be discussed in the next section.
River Transport
The products of weathering and erosion are carried to the oceans by rivers. Most of the sediment carried by the fast-flowing rivers are in suspension, as river load, while heavier particles are brought down as bed load. The carrying capacity of rivers to carry suspended sediments depends on the grain size of the sediment particles, their density and most importantly river-flow velocities. As one would expect, the amount of sediment load in a river is dependent on the area of the drainage basin (fig. 10.35).
Figure 10.35 clearly demonstrates that the average river loads decrease significantly as the area of the drainage basin increases. This may at first sound counterintuitive, however, we need to consider that an increased drainage basin area, while providing greater surface area for weathering and soil erosion to occur, also provides a larger area for sediment entrapment and/or deposition (Keller 1979). In addition, larger catchment areas are less likely to be affected by single storm events, and therefore the runoff will be less intense (Keller 1979). These are important facts to consider in the development of small islands, where changes in natural drainage patterns and/or increases in erosion are likely to occur. It is highly probable that any of these changes will have a negative impact on coral reefs in the area. In a global context, however, rivers are not the only means of sediment transport into the oceans. Table 10.3 list some of the pathways, with rough annual estimates.
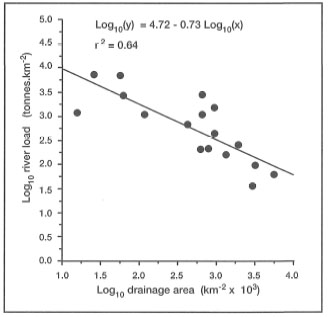
Figure 10.35. Relationship between drainage basin area and average annual sediment loads of the world's major river systems, including Solo River, East Java.
Source: Keller 1979 and Hoekstra 1989.
Sediment Origins
The composition of coastal sediments depends on a number of factors, both terrestrial and marine. One of the most obvious is the geologic history of the drainage basin and its consequent lithology and physiography (e.g., shape of drainage basin; relief and slope, orientation of streams, etc.). The amount and quality (e.g., grain size, mineralogy) of sediments from volcanic islands, sedimentary deposits, alluvial plains or raised coral islands differ dramatically as a result of their chemical and physical properties. The two most important factors that influence the rate of weathering, as well as soil erosion, are the type and amount of vegetation cover, and the frequency, intensity and duration of rainfall. Meijerink (1977) clearly demonstrate that volcanic catchment areas in Central Java produced consistently lower sediment yields than catchment areas of sedimentary origins. In all areas studied, land-use patterns and vegetation cover were comparable.
Another factor that affects the intensity of surface-water runoff is the infiltration capacity of the soils and rocks. Drainage areas characterized by well-packed, fine-grained soils and/or hard rock will most likely have higher surface-water runoff volumes than drainage areas consisting of carbonates. The major difference is infiltration capacity, which is the capacity of the soils and rocks to absorb precipitation. More porous carbonate soils have higher infiltration capacity than hard-packed clay soils, and therefore, surface-water runoff from coral islands is usually less than from comparably-sized volcanic islands. In terms of oceanic components, hydrological conditions (wave energy, coastal currents, bathymetry, seawater density, etc.) are of obvious importance.
Ambon Bay: Case Study
The influence of different lithologies and river drainage patterns on coral reef distribution can be demonstrated with islands such as Ambon, where changes occur along relatively small spatial scales. As a result of the geologic complexity, sediments along the coastline shift frequently, and this is reflected in an ever-changing mosaic of beach types. For example, while the beach deposits along the north and south coastline of Ambon Bay are rounded, pebble-sized volcanic rocks (mainly andesite), the coastline on the south and east coast of Ambon is fringed by coral reefs, and the sediments are primarily biogenic carbonates. Along the northwest coast of the island the sediments are a mixture of volcanic sands and carbonate fragments.
The steep hills in the northeastern part of Leitimur Peninsula (South Ambon) (Setayawan 1991) consist of loose Plio-Pleistocene deposits (van Bemmelen 1949). Because of the close proximity to Ambon City, the area is experiencing rapid development. Much of the original forest in the Ambon Bay catchment area has been cleared, exposing loose soils to weathering and erosion. This has resulted in severe erosion, and the once relatively clear streams are heavily polluted, and carry high loads of SPM. The coastline from Ambon City to Galala is in an accretionary phase, and the inner Ambon Bay is silting up at a rate of has resulted in severe erosion, and the once relatively clear streams are heavily pol luted, and carry high loads of SPM. The coastline from Ambon City to Galala is in an accretionary phase, and the inner Ambon Bay is silting up at a rate of 5.95 mm.yr-1 (Hermanto 1987a,b,c; Setyawan 1992). High sedimentation rates are most likely due to severe upland erosion.
Table 10.3. Annual estimates of sedimentary transport to the world's oceans.

In contrast, the mountains of Hitu Peninsula (i.e., the northern part of Ambon) are of volcanic origin (andesite, dacite and rhyolite) and are covered by extensive forests. As a result, streams along the north coast of Ambon Bay, especially in the western regions, are relatively clear. At the western part of the bay (north and south), away from anthropogenic influences, coral reefs and coral communities on non-reefal substrates are thriving.
Keeping the different lithologies in mind and looking at the distribution of reefs around Ambon, it is apparent that the most extensive coral reef development has occurred in areas where streams do not discharge large quantities of fine terrigenous sediments, which originate mostly from the erosion of unconsolidated sedimentary rocks. Inner Ambon Bay receives high volumes of river runoff during the rainy season, and as a result, only marginal reef development has occurred. Natural land runoff has been greatly altered by upland development, resulting in higher turbidity than in the past. In 1985, Sutarna (1989) recorded 41 coral species, with an average of 20 species per study site (three sites, Poka, Kate-Kate and Halong). However, percent coral cover was high, ranging from 23%-56%. During a 1992 survey of the marginal reef at Poka, the coral community consisted of 10 species (Goniopora lobata, Pontes lutea, P. lichen, Favia matthaii, Acropora humilis, Lobophyttia hemprichii, Fungia (Danafungia) danai, Fungia (Verrillofungia) concinna, Fungia (Pleuractis) scutaria, Fungia (Ctenactis) echinata), with less than 10% coral cover, compared to 16 species and 56.2% coral cover in a 1985 study (Sutarna 1989). As is the case for most of the inner Ambon Bay, the shallow reefs are being sedimented over by silt, and sandy silt (Hermanto and Suhartati 1989). In the eastern part of the inner bay, aggradation of a small mangrove stand by sand, muddy sand and mud is proceeding at a rate of about 1.2-1.6 m.yr-1 (Hermanto et al. 1989). The rapid progradation of the small delta may indicate that the bed load of the stream is large relative to its discharge volume, thus the bulk of the sediments are being deposited at the seaward edge of the delta (Wright 1989).
In comparison to the marginal reef development in Ambon Bay, extensive coral reefs are found along the southern shoreline of Leitimur Peninsula. The southern half of the peninsula is characterized by steep hills, reaching an altitude of 581 m above sea level (i.e., Mt. Horiel). Most of the rocks are volcanic in origin, consisting of andesites, pyroxenes, granites and periodites (van Bemmelen 1949). A small mini-barrier reef, 1 km in length, is located just to the north of Tanjung Hatiari about 300 m offshore. A narrow fringing reef extends along much of the southern coastline, but at Tanjung Hutumuri it widens, and a diverse coral community, dominated by tabulate acroporids, extends up to 300 m offshore. Interestingly enough, the reef displays a prominent spur-and-groove system that extends 20-50 m offshore. On the uppermost surfaces of the spurs, exposed to heavy seas, were numerous caespitose colonies of the beautiful sky-blue (azure) Acropora azurea (Tomascik, pers. obs.), not known to extend beyond Australian waters (Veron 1986). Acropora azurea has recently been found in Japan (van Woesik, pers. comm.).
Because of the local physiography, much of the southern half of Leitimur Peninsula is draining into Ambon Bay, thus the southern coastline receives very little land runoff. Similar conditions exist along the eastern and northeastern coast of Hitu Peninsula, where extensive fringing reefs have developed. The continuity of the fringing reef along the south, east and northeast coast is broken by small streams and rivulets. Even though most of the streams do not carry a heavy load of sediment year-round, and many are of only seasonal in character, they nonetheless play ah important role in coastal geomorphology, since they have a significant effect on reef development. An example of this was presented to us at Tanjung Setan, on the north coast of Hitu Peninsula.
Tanjung Setan Case Study
With assistance from Yayasan Hualopu, a local NGO in Ambon, we were fortunate to visit an enchanting cove at ominously-named Tanjung Setan (i.e., Satan's Cape). Tanjung Setan is located on the densely forested northeast coast of Hitu Peninsula, a relatively untouched region of Ambon characterized by steep volcanic hills. Mt. Salahutu reaches an altitude of 1060 m above sea level. The fringing reef along this part of the coastline narrows considerably, from about 200-300 m at Tanjung Metiela to barely 10 m wide along extensive stretches of the coastline. The reef profile is quite dramatic. Unfortunately, the reef wall west of the cove is bomb-blasted over a considerable area. To the east of the cove, the shallow subtidal reef flat was dominated by an acroporid assemblage consisting of numerous colonies of Acropora acuminata, Acropora kirstyae, Acropora millepora and Acropora tenuis. From the reef flat, the reef drops to a narrow terrace, at a depth of about 5 m, from where the reef dropped down at an angle of 75°-85° (Sutarna 1991a) to depths greater than 100 m. The almost vertical walls are densely covered by scleractinians (reef-building corals), soft corals and gorgonian sea fans (fig. 10.36).
A small clear-water stream runs out from a dense, primary, tropical rain forest, across a pure black beach consisting of fine volcanic sand, mixed with flakes of mica. The cove, about 50 m wide, is at the base of a steep mountain covered by dense rain forest. Of considerable interest was the abundance of large azure iridescent butterflies (Papillio ulysses). The cove's east and west margins consisted of a raised carbonate reef about 5-8 m in height, most likely of Pleistocene age. The beach along the cove was devoid of any carbonates, and consisted of 100% fine black volcanic sand, even though the reef was within 10 m of the beach. The slope of the shelf along the cove was about 60°. The stream's low-salinity plume was clearly visible underwater a few metres from the beach, by the shimmering effect as it mixed with seawater. However, there was a sharp transition from the volcanic silt to a carbonate reef. Thus, as Darwin suggested in 1842, the breach in this particular fringing reef is also caused by deposition of silt and not by salinity, whose effect would not have such sharp boundaries. A few experiments with the fine volcanic sand revealed that when disturbed, it immediately settled to the bottom, separating from the abundant mica flakes which had slower settling rates, producing a dramatic underwater reflective light display. Dumping handfuls of the black beach sand into the water produced similar results. The fine sand, while being almost silt size, is quite dense and sinks rapidly to the bottom. Furthermore, organic detritus in the sand must be insignificant, since disturbing the bottom sediments did not reduce the clarity of the water as expected. No shells of any kind were observed in the sediments. Therefore, the physical properties of the sediments may play a significant role in the highly localized impact, which has resulted in the breach of the fringing reef. The steepness of the slope is an additional factor for consideration.

Figure 10.36. The drop-off at Tanjung Setan, northeast Ambon (depth 25 m). Myripristis vittata (Immaculate squirrelfish) in front of a large (1.5 m high) gorgonian sea fan (possibly Plexauridae). Deep-water Acropora sp. upper left.
Photo by Tomas and Anmarie Tomascik.
Effects of Large Rivers
The effect of large rivers on the distribution of coral reefs in Indonesia seems to be relatively clear-cut. The Mahakam River (East Kalimantan), being one of the largest in Indonesia, has high discharge rates of up to 1500 m3.sec-1. With SPM concentrations reaching 80 mg/l, the river has the potential of discharging up to 3.78 x 106 metric tonnes of terrigenous sediments into the coastal waters (Dutrieux 1991). According to Eisma et al. (1989), sediment discharge from the Mahakam River may reach up to 10 x 106 tonnes.yr-1. Most of the terrigenous sediments, however, are deposited in an extensive river delta (up to 70 km wide) with an area of about 1500 km2. The intertidal regions of the delta are dominated by extensive mangrove forests and monospecific stands of Nypa fruticans. The deltaic region is an important mixing zone, where freshwater river runoff and seawater interact in a complex manner. Turbidity is high, and SPM concentrations can be as high as 160 mg/l. The mixing process is greatly influenced by a high tidal range (2.5 m). How much of the SPM is actually transported out of the delta and into adjacent regions has not been accurately estimated; however, results from the Snellius-II Expedition indicate that the sediment load in the river drops from about 25.0 mg.l-1 (lower estuary) to less than 1.0 mg.l-1 in the adjacent offshore waters. Based on seismic profile data, Eisma et al. (1989) determined that the fluvial sediments originating from the Mahakam River extend roughly halfway across the shelf, to about the 30 m isobath (40-50 km offshore). Seaward of the fluvial fan, the sea floor consists of older sediments and old coral reefs.
The net current flow in the Makassar Strait (0-200 m depth) is in a southward direction year-round, with current velocities ranging between 6-38 cm.sec-1 (depending on the season) (Wyrtki 1961). According to the known current patterns, most of the SPM leaving the Mahakam delta should be initially transported by the net current flow to the south-southwest. About 125 km south of the Mahakam delta, the northeast to southwest trending shelf turns sharply to the southeast, which may deflect the south-flowing current into a more southeasterly direction. Eisma et al. (1989) were able to identify the low-salinity (i.e., 33.0-34.0 psu) Mahakam River plume some 400 km to the southeast of the delta. The suspended particulate matter concentrations were between 0.3-0.5 mg.l-1, and the thickness of the river plume was between 50-100 m (Eisma et al. 1989). Outside the river plume, SPM concentrations dropped to less than 0.1 mg.l-1, except in the surface water. The average organic content of the SPM in the Makassar Strait is about 55%, but extremely variable (i.e., range 4% to 95%). The river plume is characterized by floes of SPM (i.e., marine snow or macroflocs) ranging between 10-100 um. Based on δ13C analysis of the organic fraction of the SPM (δ13C: -27.24 to 23.72), it seems that most of the organic matter is of autochthonous origin (phytoplankton preferentially extract 12C, leading to an enrichment of 13C; Ganssen et al. 1989). The terrestrial origin of SPM in the Mahakam delta was indicated by more negative δ13C values, which range between -28.52 to -26.50 (Eisma et al. 1989).
To what extent does the Mahakam River plume affect the distribution of coral reefs along the mainland, and in the adjacent shallow-water coastal areas, is an interesting question. The nearest coral reefs to the south of the Mahakam delta are located along the mainland coast just south of Balikpapan, about 75 km south of Mahakam. The coastal habitats between the Mahakam delta and Balikpapan have not been investigated, and it is possible that coral reefs may have developed in some areas. Our knowledge of this area is very limited, and the condition of the reefs south of Balikpapan are no exception. Based on limited data from hydrographic charts these reefs are considered as marginal. Unfortunately, they are also located in close vicinity to Balikpapan, a major industrial and oil centre in East Kalimantan. However, about 120 km due south of the Mahakam River delta, and located 75 km offshore, is the northern limit of the Great Sunda Barrier Reef, with well-developed reefs; but surprisingly very little is known about this region. Located approximately 40 km due east of the delta, at a depth of 17-25 m, are numerous shoals; however, whether coral reefs have developed in this area is not known.
In contrast to the wide area without major reef development to the south, extensive fringing reefs have developed 20 km north of the Mahakam delta, along the mainland coast just south of Tanjung Santan. From Tanjung Santan, fringing reefs extend, intermittently, in a northerly direction along a mangrove-dominated shoreline for a distance of about 50 km. The most extensive reef development occurs at Bontang Bay, 40 km north of the Mahakam, where fringing reefs and mangrove forests dominate the coastline (Nontji 1986). The largest liquefied natural gas (LNP) plant in Indonesia is located in Bontang Bay, and has been in operation since 1977. While the coral reefs are under tremendous pressure from various environmental stresses associated with the plant operation and expansion (e.g., thermal, dredging, siltation), the coral communities in 1985 were a relatively diverse group, considering the natural environmental setting. In all, there are 49 genera with 117 species, however, there is no information available on the generic and species richness prior to 1977, since an environmental impact assessment baseline study was not done.
The distribution pattern of reefs south and north of the Mahakam delta suggests that the influence of the Mahakam River may be considerable. However, the noticeable absence of coral reef development on the adjacent offshore shelf areas north and south of the delta, may be primarily due to the absence of suitable substrate. The shelf area between the delta and the northern limit of the Great Sunda Barrier Reef is characterized by flat topography, with an average depth of about 60 m. The sediments north of the barrier reef are composed almost entirely of terrigenous muds. However, it is not clear which of the three components of river discharge (i.e., salinity, turbidity or sedimentation) is the key factor impeding the development of coral reefs in the region. The rapid mixing of freshwater and seawater in the estuary and delta, greatly facilitated by the high tidal range and strong currents, make it highly unlikely that salinity has more than a local role in the general distribution pattern of coral reefs in the region.
The two key components seem to be turbidity and sedimentation. Sedimentation will have a much narrower influence than turbidity, since most SPM will settle out of the water column either in the delta or at the outer periphery. Fine silty particulate matter that significantly increases turbidity may, however, be transported by coastal currents considerable distances from the delta. The extent of the influence is obviously controlled by the discharge volume and by the regional hydrological conditions.
Land Erosion
As stressed earlier, one of the primary factors affecting sediment load in rivers is the condition of land. Vegetation provides significant protection from both the forces of erosion and weathering. Vegetation cover is also one of the easiest to alter, which, in fact, has occurred on a rather large scale, especially on Java and Bali. The main manifestation of land conversion, and subsequent mismanagement, from forests to agricultural lands or urban centres, has been a significant increase in soil erosion and increased sediment discharge into the coastal waters. This is not to say that natural erosion and sedimentation in coastal areas does not occur. For example, the fact that Jambi and Palembang are now about 50 km inland from their pre-15th century coastal position (Ongkosongo and Supriharyono 1986) is a natural phenomenon, and not a result of anthropogenic interference. Similarly, Demak in Central Java, now about 15 km from the sea, was a coastal port in the 16th century. However, the rapid horizontal growth of the Solo delta (70 m.yr1) (Hoekstra 1989), or the Wulan delta (432 m.yr-1 between 1981-1984) (Ongkosongo and Supriharyono 1986) in Central Java are clearly results of anthropogenic influences. To what extent anthropogenic impacts differ from the natural cycles of accretion and erosion is not fully quantified; however, there is one fundamental difference between the natural land runoff of the past and the present, and that is the presence of chemical substances (e.g., pesticides, herbicides, sewage and industrial pollutants) and plastics.
Land erosion has, however, become a major problem that significantly increases suspended sediment loads of many major rivers in the archipelago, and is starting to exert significant economic costs associated with mitigating measures. Recent failures of many shrimp farms (i.e., tambaks) along the north coast of Java are linked directly to the management practices, or the mismanagement, of agricultural and industrial activities in the upper-watershed areas. To what extent these activities will affect coastal ecosystems and coastal fisheries is not known, and appropriate environmental monitoring programs have not been implemented or developed.
For example, the Solo River in East Java receives about 1200 tonnes, km-2.-1 of eroded soil from its 16,000 km2 drainage basin (Hoekstra et al. 1989). The Solo River delta alone has an average deposition rate of about 17 x 106 metric tonnes per year (Hoekstra 1989). To what extent the Solo River sediment discharge influences the distribution of coral reefs along the north coast of Java and Madura has not been investigated. However, based on available data, it seems probable that the major influence will be in the eastern direction, since peak runoff occurs during the Northwest Monsoon, when the currents in the Java Sea are running in an easterly direction (Wyrtki 1961; Hoekstra et al. 1989). Information on the distribution of reefs along the north coast of Madura is not available, however, it is believed that there are none. The examination of hydrographic charts reveals that the 130-km-long northern coastline of Madura is relatively free of any significant physiographic features that can be interpreted as reefs. The 20 m isobath along the north coast of the island is about 5 km offshore, and runs parallel to the island for the entire length of the coastline. It therefore seems likely that if fringing reefs exist, the reef development should be marginal at best. Because of the unique hydrological conditions that exist during the Northwest Monsoon, the northern coastline of Madura offers some interesting research possibilities. Because of the predominant eastward transport of the river plume by the coastal currents (Hoekstra et al. 1989) during the Northwest Monsoon, it is more than likely that an environmental gradient exists along the northern coastline of Madura, in a west-to-east direction.
Sedimentation, Corals and Coral Reefs
It has long been recognized that sedimentation plays a key role in the distribution of coral communities and coral reefs (Darwin 1842). Available information from Indonesia clearly supports this general thesis. However, what is also quite apparent and needs reevaluation is the general assumption that vigorous reef development is restricted to clear oligotrophic waters. Numerous examples of extensive flourishing coral reefs in turbid coastal waters of Indonesia (e.g., Berau Islands where Secchi disk depths may be consistently less than 3 m, and where surface nutrient concentrations would be considered as evidence of eutrophication in other more oligotrophic regions) raise some important ecological, conservation and management questions.
The massive reef systems along the east coast of Kalimantan are an example where the generalization needs to be modified that reefs are "generally better-developed, have more coral species, higher coral cover, and faster rates of framework accretion the further they are from sources of runoff or the lower the sediment load in overlaying water" (Rogers 1990). However, increased sediment load in coastal areas resulting from land clearing, coral mining, dredging, construction, deforestation, drilling activities, etc., are a major threat to coral conservation and management in Indonesia. Even though many coral reef communities in the archipelago seem to be tolerant of high turbidity, it should be noted that there are no data on sedimentation for these regions, and the presence of these reefs does not imply that they will be able to tolerate additional deterioration in water quality. In fact, it is more probable that these flourishing communities may be living at the upper limits of their tolerance levels, as is often the case for many other tropical systems (Johannes 1975; Hopley et al. 1989).
The negative effects of sedimentation on coral reefs and corals per se have been recently reviewed by Rogers (1990). Based on her literature review, and considerable first-hand knowledge of the subject, Rogers (1990) suggested that sedimentation rates greater than about 10 mg.cm-2.day-1 can be considered as high or chronic. She concluded that reefs subjected to high sedimentation will have lower species richness, lower coral cover or less live coral, reduced growth rates and calcification, lower recruitment rates, decreased net coral productivity, greater abundance of branching forms, and an overall reduction in reef accretion.
Sediment Rejection by Corals
Hubbard and Pocock (1972) have demonstrated that corals have various abilities to clean themselves of sediments and other particulate matter. Indeed, it seems that some coral genera (or species), such as Fungia, for example, have adapted morphologically and physiologically to highly turbid coastal environments. Note that it is often assumed that sedimentation in turbid coastal environments is high, even though there is a serious lack of data to verify this often-used generalization. Obviously the role of currents and wave action that resuspend sediments must be taken into consideration. Local hydrological conditions and general corallum morphology are the two key factors involved in passive sediment rejections. It is, therefore, not surprising to find that branching species dominate in areas where reduced wave action and/or current velocities result in higher sedimentation rates (e.g., lagoonal environments). For example, the branching Acropora formosa is one of the most abundant lagoonal species in Indonesia.
Morphological adaptation to environmental conditions characterized by high turbidity and/or sedimentation, is an important mechanism that has enabled some groups to survive under rather marginal conditions. For example, Turbinaria peltata and Echinopora mammiformis, while being among the most efficient sediment rejectors, (Stafford-Smith and Ormond 1992), have corallum morphologies (e.g., unifacial laminae) that may facilitate passive sediment rejection. These two attributes may be the key for their apparent success on the turbid reefs in the Berau Islands, where they are among the most abundant species.
Active sediment rejection has been studied by a number of workers in the past (Marshall and Orr 1931; Hubbard and Pocock 1972; Bak and Elgershuizen 1976; Rogers 1979, 1983; Logan 1988). The results indicate that there is considerable interspecific variability in sediment rejection capabilities among the various groups. Stafford-Smith and Ormond (1992) have recently demonstrated that 42 coral species studied on the Great Barrier Reef showed some active sediment rejection capability. For example, Leptoria phrygia, a common species of clear-water upper reef slopes, is able to tolerate, without damage, sedimentation rates of up to 25 mg.cm2.day-1, but being subjected to increasing sedimentation doses from 25 to 100mg.cm-2.day-1 caused significant damage and eventual mortality (Stafford-Smith 1993). However, L. phrygia has never been observed in high-sedimentation environments (van Woesik, pers. comm).

Figure 10.37. Pontes spp. on Raburabu Reef, Berau Islands (see fig. 10.11 for location). As a result of the sheltered environment, the gently sloping reef-slope has lagoonal characteristics. Strong currents are most likely the key factor maintaining high benthic productivity in the area, as well as preventing the burial of sessile benthic organisms, such as the scleractinians.
Photo by Tomas and Anmarie Tomascik.
It appears that some coral groups that have been experimentally shown to have strong sediment rejection capabilities, are among the most common species in the more turbid lagoonal waters, where sedimentation is assumed to be high as a result of reduced currents and turbulence. For example, species with strong sediment rejection capabilities, such as Pectinia spp., Turbinaria spp. and Echinopora spp., are also abundant in many lagoonal environments (Stafford-Smith and Ormond 1992). On the other hand, some groups with strong capabilities to reject sediments (e.g., Gardineroseris planulata, Fungia repanda, LobophyIlia spp., Symphyllia spp., Favia pallida, Oulophyllia crispa, Diploastrea heliopora) may be rare in some turbid environments (Stafford-Smith and Ormond 1992), while they are common in turbid environments elsewhere (van Woesik, pers. comm.). In these species, sedimentation may not be the key factor limiting existence, since they may be more sensitive to reduced light levels which may influence their energy budget, as sediment rejection is not without energy costs. According to Stafford-Smith and Ormond (1992), Pontes spp., which is a dominant coral group in turbid coastal environments (e.g., Berau Islands), has an apparently extremely poor ability to reject fine sediments (fig. 10.37). However, Pontes have been observed in the field to slough off mucus continuously (van Woesik, pers. comm.). This process may be facilitated by strong currents. Van Woesik (pers. comm.) also points out that on the Great Barrier Reef, Montipora is the most common of all corals in turbid environments, yet it is a very poor sediment rejector.
Diploastrea heliopora, Gardineroseris planulata and Favia pallida have strong sediment rejection capabilities, and are often very abundant on the inshore turbid reefs in the Berau Islands, where underwater horizontal visibility is often less than 5 m. The area is under strong tidal influence, and as a result, tidal currents are strong. The high turbidity in the area is partly due to the close proximity of the Berau River, which discharges high volumes of runoff, but in the more offshore reefs, turbidity is mainly a function of high primary and secondary productivity. Under these conditions, zooplankton predation and suspension-feeding may provide the additional energy needed for continual sediment rejection. Tomascik and Sander (1985) have earlier suggested that some coral species may be able to use the organic fraction of SPM for growth. Therefore, it seems likely that corals living in shallow-water coastal environments rich in organic SPM, may use the particulate organic matter as an auxiliary energy source for sediment rejection. Thus, groups such as Fungia spp. may be able to tolerate higher rates of sedimentation because they may be able to capture significant amounts of additional energy from the SPM, to maintain sediment rejection activity.
Coral Recruitment
Sedimentation affects coral communities either directly through physical smothering of coral colonies or more indirectly by influencing coral recruitment and survival. Continuous recruitment of individuals is essential for the maintenance and growth of the coral community, and ultimately reef development. A number of studies have shown that sedimentation may play a significant role in coral settlement patterns and their subsequent recruitment into the various populations. It is well-known that coral planulae prefer to settle either on vertical surfaces or in crevices, and that settlement on horizontal surfaces is often avoided. The seemingly active avoidance of upper horizontal surfaces was believed to be associated with higher sediment influx, to which coral planulae may be sensitive. Hodgson (1990) demonstrated that, indeed, coral planulae avoid settling on surfaces covered with sediment. Substrate conditions under a high influx of sediment may generate a complex set of physical, chemical and/or biological cues that will inhibit planula settlement.
However, coral planulae in the water column also receive signals from their immediate environment. High concentrations of suspended particulate matter and/or changes in light intensity associated with increased turbidity may generate negative cues that are detected by the planulae, thus inhibiting their settlement until more favourable conditions are encountered. High concentrations of SPM may directly influence planulae settlement behaviour through direct physical action (i.e., abrasion), thus producing cues that delay their settlement. The reduction of light intensity may add a physiological component to the overall response through the reduction of zooxanthellae photosynthesis, which may generate specific physiological cues that delay settlement.
Suitable Substrates
Coral communities can develop on many types of substrates, providing the substrate is not mobile, most of the time. However, coral reef development requires a relatively stable substrate. In general, reef development on loose or unconsolidated substrates is inhibited or interrupted by the frequent shifting of the substrate on which the corals are attached, often resulting in the death of the colonies' However, if a suitable rubble substrate exists below the effects of wave action (i.e., below wave base), but shallow enough that hermatypic organisms can thrive, a coral reef may develop.
NUTRIENTS
Phosphorus and nitrogen are two key nutrients that are usually in shortest supply in marine environments, and therefore, it is the concentrations of these nutrients that control the extent of photosynthetic activity and primary production. However, low nutrient (phosphorus and nitrogen) concentrations are characteristic of oceanic coral reef ecosystems. They are believed to be essential for maintaining the dominance of animal-plant symbionts (e.g., reef-building corals, sponges, molluscs, etc.), crustose coralline algae, and other calcium carbonate-producing algae and invertebrates which are responsible for the build-up of the reef complex. Nonetheless, a continuous supply of inorganic nutrients is essential for the maintenance of metabolic processes and the general function of the system as a whole. While coral reefs can thrive in highly oligotrophic oceanic waters, their function is disrupted when the system is subjected to a high influx of nutrients. In fact, eutrophication (excess input of nutrients) of coastal marine ecosystems has been recognized as a serious problem worldwide.
It seems that coral reefs have suffered from excess nutrients in the past. Hallock and Schlager (1986) have suggested that the drowning of carbonate platforms in the geologic past is related to the sensitivity of scleractinian corals to nutrient enrichment of their environment. They suggested that excess nutrients will cause a reduction in water clarity, phosphate inhibition of calcification, biotic disruptions, and excess rates of bioerosion. These responses to excess nutrient subsidies may have, in the past, significantly reduced the ability of reefs to keep up with rising sea levels, and subsequently many of them drowned. High phosphate concentrations, in particular, are known to inhibit skeletal growth in reef-building corals, and to produce blooms of both phytoplankton (e.g., red tides) and benthic algae, which may grow to dominate the reef community at the expense of corals and other reef-building invertebrates.
Many of the coral reefs in eastern Indonesia are located in a region subjected to seasonal upwelling, where high nutrient concentrations support high productivity. In addition, local tidally-induced upwelling seems to play an important role in the overall productivity of the coastal waters. Thus, coral reefs in the archipelago are seldom if ever under nutrient limitation. In addition to the efficient recycling of nutrients by the reef community, and nitrogen fixation by the reef cyanobacteria and other nitrogen-fixing organisms, the reefs are continually supplied with nutrients by currents flowing over the reefs. Phosphate, in particular, does not seem to be in short supply, even though it is speculated to be the key limiting nutrient in the marine environment (Smith 1984).
During the Snellius-II Expedition (1984-1985) to the Banda Sea, phosphate (PO4) concentrations in the upper 25 m of the water column ranged between 0.10.15 μm during the Southeast Monsoon (i.e., upwelling period), and between 0.04-0.06 μm during the Northwest Monsoon (Wetsteyn et al. 1989). Nitrate concentrations (NO3) during the Southeast Monsoon range between 0.3-0.7 μm, while during the Northwest Monsoon, nitrogen seems to be limiting, since the concentrations were below instrument detection limits. Thus, there is a pronounced seasonality in nutrient availability, driven by the monsoonal upwelling system. The Banda Sea region has some of the most diverse reef communities in the archipelago, thus upwelling does not seem to have a negative influence, as would be expected. While there are no quantitative data available on seasonal fluctuation in reef productivity, based on our observations in the Banda Islands, it seems that during the Southeast Monsoon, the reef-fish biomass is considerably higher. In particular, the reef-associated pelagic species seem to be more abundant during the upwelling period. Higher benthic productivity (i.e., benthic macroalgae) during the upwelling period has not been observed. High biomass of reef herbivores may play an important role in keeping the macrophytes biomass down. However, nutrients may be a significant problem where overfishing has reduced the herbivore populations.
The extensive fringing reefs and offshore patch reefs in the western regions of the archipelago are most likely subjected to higher average nutrient concentrations as a result of their proximity to large islands. Furthermore, these reefs may also be subjected to frequent nutrient pulses associated with periodic storm-event runoff. Nutrient concentrations in these regions are seldom limiting. In the western Java Sea, surface phosphate concentrations during the Southeast Monsoon (i.e., dry season) range from 0.12-0.25 μm while during the Northwest Monsoon the concentrations range from 0.09-0.20 uM. In comparison, the inshore waters are considerably enriched with both nutrients. The [PO4] during the Northwest Monsoon in Jakarta Bay range between 0.05-2.92 μm, while during the Southeast Monsoon, the range is between 0.17-1.15 μm (Ilahude and Liasaputra 1980). In contrast to the Banda Sea, where nitrogen can be limiting, [NO3] in coastal waters of Jakarta Bay are usually high year-round. In the offshore waters of Jakarta Bay, [NOs] during the Northwest Monsoon range between 0.04-6.98 μm, and 0.05-4.01 μm during the Southeast Monsoon (Ilahude and Liasaputra 1980).
Similar conditions exist in other coastal regions not affected by large cities such as Jakarta. For example, [POJ in the Natuna Sea during the Northwest and Southeast Monsoons range between 0.06-0.40 μm and 0.07-0.85 μm, respectively. Nitrate concentrations during the Northwest and Southeast Monsoons are 0.15-3.38 μm and 0.17-1.25 μm, respectively.
Of major concern to the future integrity of many coral reefs in close proximity to cities are anthropogenic impacts associated with sewage pollution and other activities that introduce excess nutrients into the coastal environment. Sewage discharge into coastal waters results in the introduction of dissolved inorganic nutrients (i.e., NO3, PO4, NH3), dissolved organic material and suspended solids (Pastorok and Bilyard 1985). The response of coral communities to nutrient enrichment is complex. However, there is sufficient evidence to suggest that with sufficiently high doses of nutrients a major shift from an autotrophic to a heterotrophic system will occur. This has already taken place in Jakarta Bay (Tomascik et al. 1993). At lower levels of nutrient enrichment the responses are mixed; nonetheless, at some stage of nutrient enrichment, coral growth rates begin to decline, reproductive output and recruitment rates are reduced, and major changes in community structure occur (Tomascik and Sander 1985, 1987a,b; Tomascik 1991).
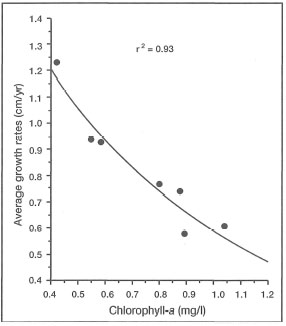
Figure 10.38. The relationship between the average growth rates (cm/yr) of Montastrea annularis and average yearly chlorophyll-a concentrations (mg/l) in Barbados, West Indies. The relationship is represented by the equation: y = -1.53[log10(x)] + 0.60; P<0.01.
Source: Tomascik and Sander 1985.
EUTROPHICATION
Bell (1993) suggested that chlorophyll-a concentrations are a good indicator of the status of eutrophication, and a threshold value of 0.5 mg.m3 was suggested for the Great Barrier Reef. This value will, however, vary between regions, since in Indonesia, for example, seasonal upwelling in the eastern regions of the archipelago can significantly increase chlorophyll-a values above 0.5 mg.m-3. Corals are able to adapt to a wide range of natural environmental conditions. Elevated nutrients and planktonic productivity associated with upwelling is not harmful to coral reef systems as long as water circulation removes excess production off the reefs and maintains water clarity. However, once productivity increases to such an extent that a significant reduction of water quality occurs, normal functioning of the coral reef system may be affected (Tomascik et al. 1993). Tomascik and Sander (1985) have earlier demonstrated a strong inverse relationship between the growth rates of Montastrea annularis, a dominant reef-building species in the Caribbean, and chlorophyll-a concentrations in the coastal waters (fig. 10.38). It was also demonstrated that increased eutrophication has a negative impact on coral recruitment rates (fig. 10.39).
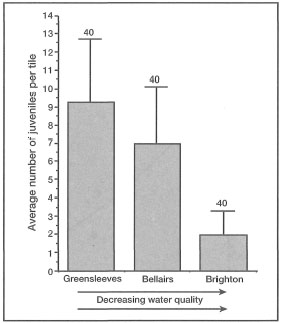
Figure 10.39. Coral settlement rates in Barbados, West Indies. Average number of juvenile corals per tile (±STD) calculated for three areas under different environmental conditions. Greensleeves fringing reef is located in an area relatively unaffected by major anthropogenic influences; Bellairs Reef is located in close proximity to a major tourism area; and Brighton Reef is located in an area polluted by urban and industrial effluents (e.g., power plant, rum refinery, deep-water harbour).
Source: Tomascik 1991.
While eutrophication is a serious problem in many areas in Indonesia (e.g., north coast of Java, Lampung Bay, Ambon Bay, south coast of Bali), appropriate coral reef monitoring studies (i.e., biotic and abiotic) have so far not been implemented. The importance of these studies cannot be overstated, since they would not only provide an early warning to potential problems, but they would provide much-needed information on the response of reefs in upwelling areas to additional nutrient subsidies.
CIRCULATION
Tides. While sea turbulence associated with atmospheric forcing is an essential factor in reef development, tides are the dominant force governing coastal, and therefore reefal, circulation patterns in the archipelago. Tidal magnitude varies from the "micro-tidal" regime of Kepulauan Seribu (Java Sea), where the tidal range is measured in centimetres (Jordan et al. 1993), to regions where the tidal range is in excess of 8 metres (i.e., Selat Muli, Irian Jaya). The most obvious effect of tides on coral reefs is the cyclic nature of subaerial exposure and inundation of extensive reef flat areas (fig. 10.40).

Figure 10.40. Extensive intertidal reef flat areas are a characteristic feature of many Indonesian reefs. The Latondu Reef is located at the northwest tip of Taka Bonarate Atoll, with a reef flat area in excess of 45 km2. The reef has two large discrete vegetated cays, Latondu Kecil and Latondu Besar (about 60 ha and 150 ha, respectively) located about 3.5 km apart.
Photo by A. Wicaksono.
Thus, tidal range has a pronounced effect on reef structure and distribution patterns. However, much less understood, but perhaps more important, especially in Indonesia, are the effects of tidally-driven coastal currents and local tidally-induced upwelling events. We have observed and dove in tidal currents with velocities in excess of 3.6 m.sec-1 (e.g., Banda Islands) (Tomascik, Mah and van Woesik, pers. obs.), while Best and Boekschoten (1988) reported tidal currents of 8.2 m.sec-1 in the narrow straits of Eastern Nusa Tenggara (e.g., between Komodo and Flores). Tidal currents of comparable velocities apparently occur in Skjerstad Fjord in northern Norway, where they have been measured at 8 m.sec-1 (Dietrich et al. 1975), as well as on the Pompey Reefs, Great Barrier Reef (van Woesik, pers. comm.). For most benthic organisms, tidal velocities of this magnitude may significantly enhance dispersal efficiency to suitable down-stream habitats (i.e., sinks). However, strong tidal regimes may also function as effective dispersal barriers and local isolating mechanisms among the numerous island groups of the archipelago (Best and Boekschoten 1988).
Oxygen Requirements. Ocean currents play an important role in maintaining a continuous supply of oxygen to reefal communities. The distribution of dissolved oxygen in coastal waters is regulated through: 1) exchange with the atmosphere, and 2) the biological processes of photosynthesis (which consumes carbon dioxide and produces oxygen) and respiration (which consumes oxygen and produces carbon dioxide). Oxygen flux from the atmosphere into surface waters is determined by the temperature and salinity of the seawater as well as by the dynamic state of the surface layer. Since tropical waters have high temperatures and salinities, the concentration of dissolved oxygen is inherently lower when compared to cooler-temperate waters.
It is widely recognized that coral reef organisms live close to their lower oxygen limits (Johannes 1975). Since high water temperatures maintain the metabolic activity of coral reef organisms at a higher rate, any activity which lowers the oxygen concentration may have a major impact on coral reef biota. It is most probable that oxygen may be an important limiting factor in coral reef areas in which water circulation is restricted. As was clearly demonstrated by Verwey (1931a), oxygen demand of the coral reef community during night hours is naturally high.
Physical Abrasion. As stated earlier, the qualitative nature of SPM in reefal waters will have considerable influence on the corals themselves. For example, large SPM particles mainly of inorganic origin (sand, volcanic ash, silt, etc.) may be highly abrasive in fast-flowing currents. High-velocity tidal currents in the archipelago are a well-known phenomenon, and together with SPM may constrain reef development in areas prone to high siltation and fast currents. For example, the south coast of West Java is exposed to the full swell of the Indian Ocean, and the tidal currents running along the coastline can reach velocities of up to 1.5 m.sec-1. Nonetheless, a well-developed fringing reef (yet to be studied) exists along most of this high energy coastline. The reef flat is only about 50-100 m at its widest, and in many areas consists mainly of Porolithon and other calcareous algae. Seaward of the breaker zone, the reef gives way to a hard substrate consisting of what seem to be volcanic rocks. The water is turbid, with horizontal visibility of about 10 m. The dominant coral in the area is Tubastrea micrantha, whose colonies are about 1 m in height. The bottom is swept by strong currents with visible sand scouring. The SPM in the water column consists predominantly of large grains of resuspended coral sand, most likely originating from the surf zone. While the area had a very sparse coral cover (<15%) and low species richness (about 20 coral species), the diversity of reef fish was surprising, especially in the Family Pomacanthidae. The species present were Chaetodontoplus mesoleucus, Apolemichthys trimaculatus, Plygoplites diacanthus, Pomacanthus xanthometopon, P. sexstriatus, P. navarchus and P. imperator.
Currents. Ocean currents are essential for transporting nutrients, larvae, and sedimentary materials. They are important for flushing and purifying wastes, and for maintaining patterns of scour and fill. Water velocity and turbulence also have a strong influence on the general morphology and taxonomic composition of coral reef ecosystems. Water circulation around and within coral reefs is very complex, and, as a result, makes the prediction of pollutant trajectories and concentrations very difficult. It is therefore essential that appropriate and well-designed physical oceanographic studies are conducted for all development activities near coral reef areas. Human activities such as harbour construction, channel dredging or coral rock mining may produce far-reaching biological effects, especially downstream of the activities. Altered circulation may produce conditions unsuitable for new recolonization of reef biota, thus inhibiting possible recovery.