Chapter Six
INTRODUCTION
Prior to 1911, very little was known about the nature or the general distribution of coral reefs within the Indonesian Archipelago. The earliest synthesis of information on the distribution and the type of coral reefs in the archipelago appeared in Darwin's 1842 monumental work on the structure and evolution of coral reefs, which also included the first world distribution map of these systems. Darwin (1842) in his classification of coral reefs distinguished three main types, fringing reefs, barrier reefs and atolls. However, the structural characteristics of reef forms (e.g., fringing and barrier reefs) on which Darwin based his broad and simple classification system were only loosely defined (Stoddart 1975). As coral reef research, especially coral reef geomorphology, gathered momentum to address the "coral reef problem" (Davis 1928), numerous new structural terms, considered as intermediate and end-members of Darwin's original sequence, were added to the classification (Davis 1928; Stoddart 1975). From a worldwide perspective, recent additions of new coral reefs to Darwin's original distribution map can be considered as relatively minor when compared to the major transformation of reef distribution patterns in the Indonesian Archipelago.
Early coral reef distribution maps were based largely on the interpretation of available hydrographic charts (Darwin 1842; Niermeyer 1911; Joubin 1912; Molengraaff 1929), and as a result, the distribution of coral reefs in the remote regions of the vast Indonesian Archipelago was not well-known; indeed, there were serious doubts whether barrier reefs and atolls existed in the archipelago at all (Molengraaff 1929). This lack of knowledge was clearly reflected in Darwin's 1842 world distribution map for coral reefs. Indeed, one of the most interesting features of Darwin's original map was the total absence of atolls and barrier reefs in the archipelago, with fringing reefs dominating the southern and north-western fringes, but the inner archipelagic sea, especially around Sulawesi, relatively devoid of coral reefs. However, Darwin's rather detailed account, considering the limited amount of information available to him at the time, of coral reefs in the then East-Indian Archipelago, was the first synthesis of coral reef distribution for this region. While Niermeyer (1911) is generally credited as being the first to call attention to the existence of atolls and barrier reefs in the Indonesian Archipelago (Molengraaff 1929), it seems that Darwin (1842) was well aware of this fact as well. Describing the now rather well-known atolls and barrier reefs of the Tukang Besi island chain in Southeast Sulawesi, Darwin wrote:
From the imperfect charts I have seen, I have not been able to decide whether they belong to the atoll or barrier-class.... I think it, however, exceedingly probable that some of them ought to be blue [atolls].—DARWIN 1842
It appears that Darwin's classification of Indonesian coral reefs was influenced by two factors. Firstly, he did not have access to the detailed hydrographic charts that only later became available, and secondly, his classification of Indonesian reefs, based on reports from earlier voyages, seemed to have been influenced by his knowledge of many raised coral islands in the archipelago. This may have been a key factor, since Darwin's theory of atoll and barrier reef formation is based on subsidence of the underlying basement. Darwin's exclusion of barrier reefs and atolls from the Indonesian Archipelago was, however, taken at face value, which resulted in a widely held assumption, at the time, that barrier reefs and atolls did not occur in the archipelago (Molengraaff 1929). It seems clear, however, that Darwin's hesitancy to classify many reefs in the Indonesian Archipelago as either barrier reefs or atolls was mainly a result of insufficient data.
From the distance to which they extend from the land where the sea is very deep, I am strongly inclined to believe they ought to come within the barrier class, and be coloured blue; but I have been forced to leave them uncoloured.—DARWIN 1842
It is interesting to point out that the absence of barrier reefs and atolls in the archipelago was also a result of the difficulty in observing these formations (fig. 6.1). Why this may be so was clearly elaborated on by A.R. Wallace, when observing, for the first time in Indonesia, a true barrier reef at the Watubela (Matabello) Islands in the eastern Banda Sea:
In a sheltered archipelago they will seldom be distinguishable, from the absence of those huge rolling waves and breakers which in the wide ocean throw up a barrier of broken coral far above the usual high-water mark, while here they rarely rise to the surface.—WALLACE 1869
Nowhere is this statement more appropriate than in the sheltered Gulf of Tomini, where an extensive barrier reef system surrounds most of the Togian Islands. The distribution of coral reefs in the central and southeastern regions of the archipelago (e.g., Flores Sea, Molucca Sea, Banda Sea, Sulawesi Sea) remained relatively unknown until the publication of new hydrographic charts by the Dutch navy in the early 1900s. With the exception of the Indonesian Archipelago, Darwin's map has proved to be quite accurate.
Niermeyer (1911) was the first to 'formally' review the general distribution patterns and types of coral reefs in the Indonesian Archipelago. With the aid of new hydrographic charts that became available from the Royal Dutch Navy, Niermeyer (1911) was able to demonstrate conclusively, for the first time, the presence of numerous true barrier reefs and atolls, the existence of which were earlier doubted, but alluded to by Darwin (1842). Almost 20 years later, based on the work done by Escher (1920) and aided by the availability of additional new and upgraded Dutch hydrographic charts, Molengraaff (1928) provided the first comprehensive treatment on the general distribution and development of coral reefs in the archipelago, while Umbgrove (1928) gave the first synthesis of the geomorphology of coral reefs in Jakarta Bay and Pulau Seribu. This work was later expanded upon by Umbgrove (1947). Coral reef science, as such, received a significant boost during the highly successful Snellius Expedition (1929-1930). The reports generated by the expedition provided a wealth of new information on the general distribution and nature of coral reefs in the eastern parts of the archipelago. More significant, however, was the expedition's contribution to physical oceanography and marine geology, especially in the field of coral reef geomorphology.

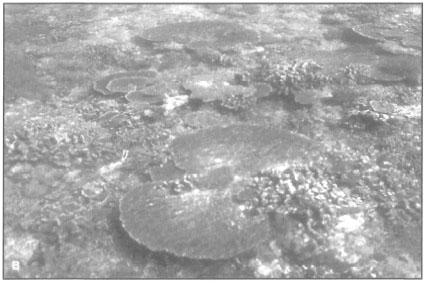
Figure 6.1. Among the most interesting features of the Indonesian Archipelago are the calm intra-archipelagic seas with numerous coral islands fringed by palm-lined white sandy beaches. A) The calm waters around Maringki Island, Banggai Archipelago, hide all traces of the barrier reef that runs along the southeast margin of the archipelago. The well-known surf zone (i.e., the breakers) of the classical barrier reefs, such as Bora Bora, are a rare sight in the archipelago. B) The transition from deep oceanic water to shallow reef crest is without warning. Presence of large tabulate acroporids, characteristic of high-energy seaward reef zones, is most likely due to strong tidal currents rather than to wave action. Photo taken about 1 km off the coast of Maringki Island.
Photos by Tomas and Anmarie Tomascik.
THEORIES OF CORAL REEF ORIGINS
Coral reefs were well-known features of the tropical seas long before they became a subject of scientific study. Early sea voyagers that ventured into the vast tropical seas generally referred to these navigational hazards as atolls (lagoon-islands), barrier reefs (encircling reefs) and fringing reefs (shore reefs) (Darwin 1842). Coral reefs became a subject of scientific interest during the early 19th century, when new information on their nature, geomorphology and distribution became available from a number of expeditions of discovery and scientific collecting. For example, Von Chamisso (1821) observed that the upward-growing reef-building corals seem to do best in the shallow surf zone at the outer periphery of the reef. This led him to speculate that the seaward margins of reefs growing upwards from their foundation would reach the surface first, thus forming the characteristic annular shape of atolls. Quoy and Gimard (1825), who participated in two round-the-world expeditions, observed that vigorous growth of reef-building corals is restricted mainly to shallow-water habitats. Based on their observations and collections during two expeditions, Quoy and Gimard (1825) were of the opinion that the annular shape of atolls is a result of upward growth of reefs that became established around the rims of extinct and submerged volcanic craters. Lyell (1832) was one of the strongest supporters of the volcanic crater theory, which, however, he later dropped in favour of Darwin's theory of subsidence. It is interesting to point out that Darwin (1842) did not totally reject the submarine crater theory, for he wrote: "… if the rim of a crater afforded a basis at the proper depth, I am far from denying that a reef like a perfectly characterized atoll might not be formed …". The flourishing coral reef communities that we have observed on the slopes of an active submarine volcano Banua Wuhu, Sangihe Islands, North Sulawesi, suggest that coral reef development on submarine volcanic craters may indeed be possible, even on active volcanoes. Whether atolls are formed in this fashion remains unknown, but it is highly unlikely. While growth of corals on tops of submerged extinct, or active, volcanic craters is possible, the submarine crater theory'cannot explain the great depths from which the reefs seem to originate, the various shapes and sizes of atolls, nor the similarity in lagoonal depths which would require a large number of volcanic craters to.be at the same depth (Darwin 1842). Another theory mentioned, but rejected, by Darwin (1842) was that atolls may be formed by a reef growing on a leveled, detached bank. In fact, he suggested that this may occur in the West Indies (Darwin 1842, p. 89).
While a number of theories were proposed to explain the formation and general features of oceanic atolls, barrier reefs were generally neglected. Based on their observations of steep cliffs along much of Tahiti's coastline, Tyerman and Bennet (1832) suggested that the extensive barrier reefs surrounding the steep volcanic island have formed at the seaward edges of wave-cut platforms. It is interesting to point out that while observing the same coastline in Tahiti seven years later, Dana (1872) arrived at a fundamentally different conclusion. Dana (1872) saw the coastline of Tahiti as embayed valleys and used this feature as evidence of subsidence, thus providing new support for Darwin's theory. Hopley (1982) provides a detailed historical review of theories accounting for the formation of the Great Barrier Reef.
Darwin's Theory of Subsidence
Several theories have been advanced to explain the origin of atolls or lagoon-islands, but scarcely one to account for barrier-reefs. From the limited deptiis at which reef-building polypifers can flourish, taken into consideration with certain other circumstances, we are compelled to conclude, as it will be seen, that both in atolls and barrier-reefs, the foundation on which the coral was primarily attached, has subsided; and that during this downward movement, die reefs have grown upwards.—DARWIN 1842
The above statement summarizes one of the most fundamental geomorphological theories in coral reef science. Based on his observations and collections made during the voyage of the Beagle, Darwin formally introduced his elegant solution for the formation of atolls through slow subsidence and upward growth of corals to the Geological Society (London) in 1838 (Darwin 1838). Four years later, Darwin (1842) presented his work on coral reefs in his influential publication, The Structure and Distribution of Coral Reefs. Almost 150 years later, one of the coral reefs' most respected geomorphologists had this to say about Darwin's 1842 work: "In many respects, it would be wise for modern scientists to read Darwin's book carefully, for they would find many facts and opinions which are often presented as recent discoveries…" (Guilcher 1988).
It should be made clear, however, that Darwin's genetic theory of atoll formation by subsidence relates mainly to the development of oceanic atolls which have evolved under very different conditions from many other reef types found in geologically young and tectonically active areas (Hopley 1982). The ensuing controversy on the origin of barrier reefs and atolls that followed Darwin's 1842 publication lasted for over a hundred years, and was mainly a result of attempting to solve a complex problem requiring more than one explanation with a single solution. As Hopley (1982) pointed out, all theories of atoll formation attempted to provide a single evolutionary sequence to address three basic problems that were the focal point of the controversy, namely: 1) that the growth of reef-building corals is restricted to the euphotic zone; 2) that most reefs originate from depths well below the euphotic zone; and 3) that atolls exhibit great similarity, especially in the nature and depth of their lagoons.
Darwin's theory of barrier reef and atoll formation through subsidence and upward growth of coral was an impressive feat of deductive logic. It has often been implied that Darwin formulated his theory after seeing barrier reefs and atolls during his voyage on the Beagle (1832-1836). However, as Stoddart (1962b) pointed out, Darwin conceived the idea that fringing reefs, barrier reefs and atolls were genetically related through slow subsidence long before he saw his first Pacific atoll. The concept of the subsidence theory was formulated during the voyage of the Beagle, while Darwin was studying uplifted coastal formations on the west coast of South America. The presence of marine fossils high up in the Andes Mountains indicated to him that vertical movements of the earth's crust must occur in many areas.
No other work of mine was begun in so deductive a spirit as this: for the whole theory was thought out on the west coast of South America before I had seen a true coral reef. I had therefore only to verify and extend my views by a careful examination of living reefs. But it should be observed that I had during the two previous years been incessantly attending to the effects on the shores of South America of the intermittent elevation of land, together with the denudation and deposition of sediment. This necessarily led me to reflect much on die effects of subsidence, and it was easy to replace in imagination the continued deposition of sediment by the upward growth of corals. To do this was to form my theory of the formation of barrier reefs and atolls.—IN BARLOW 1958
During the homeward leg of the Beagle, Darwin visited Tahiti, where he was able to make a number of field observations that provided the necessary support for his subsidence theory. Based on his observations of the barrier reef and lagoon surrounding Moorea, Darwin concluded that barrier reefs and atolls are not very different in terms of their geomorphology. Darwin's view of Moorea from neighboring Tahiti made a lasting impression which he recalled when he wrote:
I was much struck with this fact, when viewing, from the heights of Tahiti, the distant island of Eimeo [Moorea] standing within smooth water, and encircled by a ring of snow-white breakers. Remove the central land, and an annular reef like that of an atoll in an early stage of its formation is left; remove it from Bolabola [Bora Bora], and there remains a circle of linear coral-islets, crowned with tall cocoa-nut trees, like one of the many atolls scattered over the Pacific and Indian Oceans.—DARWIN 1842
Darwin's theory of atoll formation through subsidence is illustrated in figure 6.2. The theory requires that the slow subsidence of the foundation (e.g., volcanic island) is matched by the upgrowth of the coral.
Figure 6.3 is a simplified schematic representation of Darwin's theory, clearly illustrating the genetic nature of the theory, in which the fringing, barrier and atoll reefs are linked together in a single evolutionary sequence (Hopley 1982). The initial formation of barrier reefs and atolls begins when a volcanic island, or another suitable foundation, is colonized by scleractinian corals. It is implicit that the foundations upon which reef growth is initiated are located in environments suitable for coral reef development. If environmental (biotic and abiotic) conditions continue to be favorable, a shallow-water fringing reef will develop along much of the coastline. In time, the fringing reef will reach the surface and will begin to expand horizontally in a seaward direction. In instances where the foundation is geologically and tectonically stable, the fringing reef may expand laterally over its own seaward talus slope. While there is some evidence from recent Indonesian reefs, which, for the past 4000 years or so, had to cope with gradual sea-level drop, that this may occur on gently sloping island shelves (T. Tomascik, pers. obs.), the general concept that coral reefs grow laterally over their own tali has recently been criticized as basically unsubstantiated (Rosen 1992). Whether or not the reef grows laterally is, however, not an issue if we assume subsidence of the foundation. Once the foundation begins to slowly subside, the upward growth of coral will keep pace with the apparent sea-level rise and the fringing reef will continue to grow both vertically and laterally (i.e., relative to the subsiding island). With continued subsidence
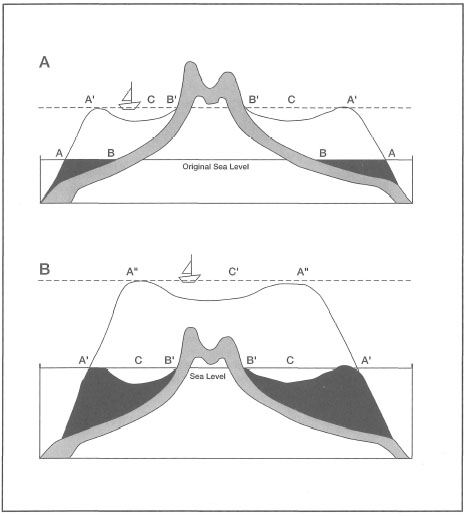
Figure 6.2. The development of barrier reefs and atolls through subsidence. Figures redrawn from Darwin (1842; No. 4 and 5). Captions by Darwin: A) AA - Outer edge of the reef at the level of the sea. BB - Shores of the island. A'A' - Outer edge of the reef, after its upward growth during a period of subsidence. CC - The lagoon-channel between the reef and the shore of the now-encircled land. B'B' - The shore of the encircled island. N.B. In this, and the following woodcut, the subsidence of the land could only be represented by an apparent rise in the level of the sea. B) A'A' - Outer edges of the barrier reef at the level of the sea. The cocoa-nut trees represent coral-islets on the reef. CC - The lagoon channel. B'B' - The shore of the island, generally formed of low alluvial land and of coral detritus from the lagoon-channel. A"A" - The outer edge of the reef now forming an atoll. C - The lagoon of the newly formed atoll. According to the scale, the depth of the lagoon and of the lagoon-channel is exaggerated.

Figure 6.3. The theoretical evolutionary sequence of barrier reef and atoll formation through subsidence and vertical growth of coral. Not to scale. A) Development of fringing reefs around a volcanic foundation. B) Slow subsidence of the volcanic foundation results in the formation of a barrier reef separated from the coastline by a lagoon. C) The submergence of the volcanic foundation eventually leads to the formation of an atoll with a deep central lagoon. Numerous coral cays may be formed within the lagoon as a result of reef growth. and complex interaction of biotic and abiotic factors, a channel will initially form between the seaward edge of the reef and the shoreline of the subsiding island. In time, the channel will widen and deepen to become a lagoon. At what stage the fringing reef becomes a barrier reef is debatable, since it generally depends on the depth of the lagoon. Sheppard (1983) suggests that a fringing reef becomes a barrier reef once the channel or lagoon is wide and deep enough to be able to take large ships. In Indonesia, the depths of barrier reef lagoons range from a minimum of about 5 m to more than 100 m. With almost continual subsidence of the foundation, only occasionally intermitted by periods of stability or even uplift, the island will eventually submerge. Once the volcanic island is submerged, leaving only an open lagoon, the annular reef becomes an atoll. Note that the lagoons of many atolls in the Indonesian Archipelago (e.g., Taka Bone Rate Atoll, Flores Sea) are dotted by numerous sand cays (i.e., vegetated low islands), whose origins are, however, reefal.
Darwin's publication of The Structure and Distribution of Coral Reefs in 1842 may be viewed as the birth of coral reef science, since subsequent to its release major research efforts were mounted to either prove or to disprove his theory. It is a remarkable fact that the coral reef controversy lasted for over a 100 years, mainly because of technological limitations. To prove the basic premise of Darwin's theory required only to drill into atolls to verify that the underlying foundations of atolls are volcanic in origin. Indeed, Darwin himself suggested such a project, for in 1881 he wrote:
I wish some doubly rich millionaire would take it into his head to have borings made in some of die Pacific and Indian Ocean atolls and bring home cores for slicing from a depth of 500 to 600 feet.—IN HOPLEY 1982, P. 4
Determining the geologic nature of the substrate beneath oceanic atolls, as well as the origin of their annular morphology, became a hotly debated topic in coral reef science. Increased criticism of Darwin's theory in the latter part of the 19th century, mainly in favour of pre-existing platform theory (Murray 1880), prompted Dana (1885) to mount a strong defense of the subsidence theory based on his observations of embayed shorelines on islands surrounded by barrier reefs. However, the debate was far from over. In an early attempt to resolve the so-called Coral ReefProblem (i.e., was Darwin correct?), the Royal Society (London) in 1896-1898 Carried out an ambitious drilling project at Funafuti Atoll, Ellice Islands (now Tuvalu) (i.e., Funafuti Coral Reef Boring Expedition). In 1898 the Funafuti boring reached a depth of 340 m; however the findings proved inconclusive, since the boring did not reach the foundation. While the upper 194 m consisted of coral limestone, the lower section of the core was entirely dolomite, possibly from a fore-reef talus (Hopley 1982). Following the unsuccessful attempt at Funafuti Atoll, The Royal Geographical Society (Australia) conducted two drilling projects on the Great Barrier Reef. The first boring in 1926 was carried out at Michaelmas Cay and penetrated to a depth of 183 m, while the second boring in 1937 at Heron Island reached a depth of 223 m (Fagerstrom 1987). Unfortunately, neither borings reached the underlying foundation, and the Coral Reef Problem remained unresolved, even though Davis (1928) provided strong circumstantial evidence in support of Darwin's theory.
In the early 1940s, as part of their oil exploration activities in the Indonesian Archipelago, the Dutch oil company Bataafsche Petroleum Maatschappij carried out two borings on Maratua Island, an uplifted atoll just off the east coast of East Kalimantan (Borneo). Kuenen (1947) analyzed the logs of the two deep cores and concluded that subsidence of at least 500 m occurred, thus providing new evidence for Darwin's subsidence theory. However, while drilling proceeded to 261 and 429 m, neither boring penetrated the bedrock and, therefore, the effort was proved once again inconclusive with regards to solving the Coral ReefProblem. Note, however, that the two borings were not conducted to address the coral reef problem. Nonetheless, based on his interpretation of the cores as well as his detailed field observation on Maratua during the Snellius Expedition (1929-1930), Kuenen (1947) provided strong support for Darwin's subsidence theory.
At about the same time (i.e., 1947) that Kuenen published his Maratua results, two deep borings were conducted at Bikini Atoll. The deepest core reached 780 m, but once again failed to reach the basement rock. The first conclusive proof of Darwin's subsidence theory came finally in 1951, when a 1340-m-deep bore at Enewetak Atoll (Marshall Islands) struck the underlying volcanic foundation (Ladd et al. 1953; Ladd 1973). Thus, it took over 100 years to prove the basic premise of Darwin's theory, and validate his subsidence theory. In subsequent drillings, Ladd and Schlanger (1960) demonstrated that the reefal deposits immediately above the volcanic basement of Enewetak Atoll (depth of 1283 and 1408 m) were of Eocene age, and contained scleractinian corals restricted to the euphotic zone (i.e., shallow-water reef-builders). Similar results (i.e., penetration of basaltic basement) were obtained from drillings at Midway Atoll (Hawaii) (Ladd et al. 1970) and at Mururoa Atoll, Tuamotus Archipelago (Aissaoui and Purser 1985; Buigues 1985; Gachon and Buigues 1985; Ruzie and Gachon 1985). Mururoa Atoll is in fact one of the most drilled atolls in the Pacific Ocean, with 13 boreholes deeper than 175 m (Bardintzeff et al. 1985). Since the boreholes were sunk in different parts of the atoll, six in the lagoon and seven in the outer rim, Mururoa Atoll remains the only atoll where the three stages of reef evolution (fringing reef to barrier reef to atoll) are known with some certainty. The extensive borings at Mururoa proved to be invaluable. Not only were Enewetak findings confirmed, but the boreholes sunk in the lagoon and the rim of Mururoa Atoll allowed a comparison of lagoonal and rim deposits, which provided the first conclusive evidence that the rim of the atoll was formed by a vertical reef build-up around a slowly aggrading central lagoon (Buigues 1985), as was originally proposed by Darwin (1842). The Mururoa and Midway cores also demonstrated that incipient fringing reefs formed while the volcanoes were still quite active. Interestingly, an excellent example of rapid reef development on still-active young volcanoes can be found at Banda Api, Banda Islands. The release of large quantities of andesitic lava during the 1988 eruption of Banda Api smothered a large section of a pre-existing fringing reef. Surprisingly, a flourishing coral community, characterized by high species diversity and high-percentage coral cover, became established on the submerged lava flow within 5 years of the eruption (Tomascik et al. in press). Guilcher (1988), in his review of the history of coral reef drilling projects in the Pacific, concluded:
These results demonstrate, with a certainty rarely obtained in geomorphology, that the seven atolls were built by corals and associated organisms during a long-continued subsidence, over a differing period in each case. In other words, the subsidence theory, proposed by Darwin and developed by Dana, Davis and others, is thus fully supported.—GUILCHER 1988
Challenges to Darwin's Theory
As is often the case in science, a number of alternate theories were proposed to challenge Darwin's theory of subsidence. However, as Hopley (1982) pointed out, the main opposition to subsidence was:"… often by zoologists who, in the view of many Darwin supporters, did not fully appreciate the geological aspect of the question". It is, therefore, not surprising that most of the theories that challenged Darwin's subsidence model have been largely discarded. Among the strongest supporters of Darwin's subsidence theory was W.M. Davis, who, in reviewing the various alternative hypotheses, concluded that:
…they all have two characteristics in common; namely defective deduction of consequences and hence a very incomplete confrontation of consequences with facts. Perhaps the chief reason for these deficiencies is that nearly all the inventors of these theories had been trained more in die biological than in geological science…. Their theories cannot be accepted because the consequences are strongly contradicted by the facts.—DAVIS 1928
Nonetheless, three hypotheses have had a significant influence on the contemporary views of coral reef origins, especially of barrier reefs and atolls. These were the glacial-control theory (Daly 1915), the antecedent-platform theory (Hoffmeister and Ladd 1944) and the karstic-saucer theory (McNeil 1954; Purdy 1974a).
The Glacial-Control Hypothesis. Sea-level fluctuations relative to the land have a number of causes and can be either local, regional or global. In the Indonesian Archipelago, local sea-level fluctuations consist of transgressions and regressions caused primarily by tectonic processes. While many areas in the eastern parts of the archipelago (e.g., Alor, Sumba, Sula Spur) have been experiencing significant uplift (i.e., relative drop in sea level or regression) during the Quaternary (Hantoro et al. 1994), some areas in the western regions (e.g., Sunda Strait) have experienced transgression (i.e., subsidence or relative rise in sea level) (Hantoro 1995a). In contrast, global eustatic (Suess 1888) sea-level fluctuations produce concurrent transgressions and regressions on the shelves of all continents, and are caused primarily by changes in the balance between glacial ice and water, as well as changes in depth of ocean basins and the buoyancy of continents.
The idea that sea levels have fluctuated widely up and down during Earth's long geologic history is not new. In fact, Aristode (c. 335 B.C.) contemplated the idea long before Agassiz (1837,1840) advanced his ideas of the "ice "age", and McLaren (1842) deduced that glaciation would lower global sea levels by as much as 120 m.
And if in some places the sea recedes while in others it encroaches, then evidently in the same parts of the earth as a whole are not always sea, nor always mainland, but in process of time all change.—ARISTOTLE METEOROLOGICA (IN LEE 1952)
While much has been learned about glacial processes, the main question of what causes ice ages is still open to discussion. Interestingly, glaciations are a relatively rare natural phenomenon, with only four or five known periods during Earth's history, each lasting only a few million years (Spencer 1972). However, sea-level fluctuations are a common feature of Earth's history (Hallam 1992). The earth has been in a glacial mode since about the middle Miocene (Veron 1995), and for the past 3 million years large quantities of ice, as polar ice-caps and continental glaciers, have covered large areas of the planet. During the Pleistocene, there were at least 17 cycles of transgression and regression (Kukla 1977; Grigg and Epp 1989), with the last major advance culminating at about 18,000 years ago. At the peak of the last ice age, about 30% of continental area was covered by glaciers and about 8% of water was in the form of ice (Spencer 1972). With regards to the history of coral reef development, it seems that sea-level fluctuations resulting from high-frequency glacial cycles, mainly under the influence of planetary processes (Milankovich 1938), may have had the most impact. Indeed, the last glacial maximum (i.e., 18,000 years ago) followed closely a strong insolation cold peak that occurred about 4000 years earlier (i.e., 22,000 years ago). The insolation cold peak resulted from changes in the Earth's angle of the elliptic as well as in the angle of Earth's axis (i.e., Earth's wobble). ,
Before any of the above information was available to coral reef scientists, Penck (1894, 1896) was among the first to link sea-level fluctuations to reef development. However, it was Daly (1915) who first fully appreciated the impact that glaciation may have on coral reef development, both in terms of sea-level fluctuations and changes in seawater temperatures. Daly's glacial-control theory became the first serious challenge to Darwin's subsidence theory, as the primary explanation for the formation of barrier reefs and atolls. The glacial-control theory (fig. 6.4) found many early supporters, notably in Vaughan (1914, 1919, 1923); however, some of the strongest opposition came from Dutch scientists working in the Indonesian Archipelago, namely Kuenen (1933a, 1947) and Umbgrove (1930, 1939c, 1947). In fairness, it should be pointed out that both Kuenen (1947) and Umbgrove (1947) agreed with many aspects of Daly's theory.

Figure 6.4. Simplified representation of Daly's glacial-control theory. A) Atoll formed during the last interglacial. B) Destroyed, truncated reef at glacial maximum. All reefs are assumed to be planed off. C) New reef develops on the truncated platform during subsequent transgression.
Simplified from Daly 1915.
These statements [i.e., referring to Ladd and Hoffmeister's (1936) critique of glacial-control theory] do not mean, however, that Pleistocene changes of sea level are of no importance to coral-reef problems. On the contrary in any reef region that factor is to be considered, but must not be overrated.—UMBGROVE 1947
Daly formulated his glacial hypothesis to explain the apparent uniformity of lagoon depths of most large barrier reefs and atolls (i.e., 70-80 m) (Guilcher 1988). However, the theory was generally taken as a new theory on the origin of barrier reefs and atolls, and therefore viewed as contradicting Darwin's theory of subsidence. Daly (1915) proposed that during the last glacial maximum global sea levels may have been as much as 120 m below present-day values, thus exposing large areas of formerly submerged foundations to rapid subaerial erosion and weathering. Combined with decreases in sea surface temperatures during the glacial maximum and subsequent increases in turbidity and sedimentation due to rapid erosion of subaerially exposed foundations, most reef-building corals were killed, especially in marginal environments (i.e., high latitudes). The destruction of reef-building corals as a result of a drop in sea surface temperatures is an important component of the glacial-control theory, since rapid erosion of exposed foundations could occur only if reefs have lost their biogenic protection at their seaward margins and reef flat areas. Once this protection was lost, exposed reefs and other foundations were subjected to rapid erosion, which resulted in increased turbidity and sedimentation, preventing further reef build-up. Without their coral reef protection, Daly (1915) visualized that most foundations, including old coral reefs as well as deeply weathered old volcanic islands, were planed off to a depth of about 70-80 m. Younger volcanic islands, not susceptible to rapid weathering and erosion, were not planed off, but rather narrow peripheral erosional platforms were formed. As sea surface temperatures began to increase and the sea levels started to rise during the interglacial, the abraded platforms were once again recolonized by corals which survived the glacial period in their Pleistocene sanctuaries. Because of ecological factors, coral reef development was assumed to be fastest at the seaward edge of the platforms. Thus, as the sea levels continued to rise and coral growth kept pace with the rising sea levels, barrier reefs and atolls were eventually formed.
Indonesian Reefs and the Glacial-Control Theory. The glacial-control theory explained a number of surface features of barrier reefs and atolls that were difficult to explain by Darwin's subsidence alone. More importantly, however, the theory linked the observed submergence of foundations with the Pleistocene meltdown of continental and polar glaciers. As Bloom (1974) commented, the role of glacial control in coral reef geomorphology is important, but not in the sense that R.A. Daly visualized. Sheppard (1983) said it well when he stated that: "At first, some regarded this as a contradiction of Darwin's theory, but in fact Darwin explained how reefs originated and Daly explained how sea-level changes and erosion could cause surface features".
Davis (1928), based on his extensive travels throughout the coral seas, offered a very strong argument against the glacial-control theory as an alternative to Darwin's theory of subsidence. In subsequent years, as more information became available, especially from reefs of the Indonesian Archipelago, major shortcomings of the glacial-control theory became evident. Nonetheless, information from other regions of tropical seas seem to have supported some of Daly's arguments (Nugent 1946). Major weaknesses of the glacial-control theory were Daly's overestimates of erosional rates, his emphasis on the detrimental nature of increased turbidity and sedimentation, and the fact that lagoonal depths of barrier reefs and atolls are not as uniform as was believed (Guilcher 1988).
Based on his extensive field work during the Snellius Expedition, Kuenen (1933a) provided the first detailed description of the geomorphology of many coral reefs in the eastern parts of the archipelago. The geological results of the Snellius Expedition provided a wealth of new information on the development of coral reefs in one of the most tectonically active regions of the world. Kuenen's studies provided compelling evidence against Daly's (1915) glacial-control theory as the only, or the main, explanation for the formation of atolls and barrier reefs, well before the borings at Enewetak Atoll conclusively proved Darwin's theory of subsidence. According to Kuenen (1933a), who studied the general geomorphology of the barrier reefs, atolls and islands in the Tukang Besi island chain in great detail: "The most convincing fact in favour of Darwin's theory is the great steepness of the submarine slopes of the atolls". The atolls of the Tukang Besi group are arranged in a linear fashion according to the tectonic structure of the substratum (Umbgrove 1947). The atolls are interspersed with rows of elevated islands, many of which are fringed by small barrier reefs. The uplifted islands consist of soft friable marls coated with reef rock (Kuenen 1947). Umbgrove (1947) observed that atolls in the southwest region of the complex are followed by a row of elevated islands, which is followed by another series of atolls and a last row of islands with elevated reef terraces. Kuenen (1947) pointed out that the submarine slopes of the atolls are much steeper (30° to 50° to a depth of 500-600 m) than the slopes of the elevated islands. He noted that while islands with fringing reefs have slopes of 30° to 50° to a depth of about 200 m, which he attributed to vertical growth of the reef, the slopes below the 200 m depth are only 10° to 20°. This led him to conclude that the atoll foundations are of an entirely different origin than those of the elevated islands. Based on his field observations of numerous barrier reefs and atolls in the tectonically-active eastern parts of the archipelago, Kuenen (1933a) rejected the glacial-control theory as a dominant force in the development of Indonesian reefs, and stated that: "In the East Indies the influence [of lower sea levels] can only have been to modify the already existing atolls to a small extent".
Based on paleontological arguments, Kuenen also rejected the assertion, as did later Umbgrove (1946c, 1947), that lower sea surface temperatures, in conjunction with higher turbidities, during the glacial maximum had a significant influence on coral survival and reef development in the archipelago. This view was reiterated by Bloom (1974), who suggested that the central Pacific and many other coral reef provinces were neither too cold nor turbid for reef development, even during glacial maxima. However, Fleminger (1986) suggested that as a result of intensified coastal upwelling, which developed as more shelf areas became exposed during Pleistocene glacial periods, sea surface temperatures in the eastern regions of the Indonesian Archipelago, roughly between Sulawesi and Irian Jaya (i.e., Wallacea), may have been much lower than is usually presumed. Fleminger (1986) argues that seasonal coastal upwelling in the northern regions of Wallacea (e.g., Sulawesi Sea, Molucca Sea, Seram Sea) during the Northwest Monsoon (i.e., Northern Hemisphere's winter) may have reduced sea surface paleotemperatures to as low as 21 °C. A similar scenario was suggested for the southern regions of Wallacea (e.g., Flores Sea, south and east Banda Sea), where strong upwelling during the Southeast Monsoon (i.e., Southern Hemisphere's winter) may have lowered sea surface paleotemperatures to as low as 20°C. According to Fleminger (1986), low sea surface paleotemperatures may have eliminated many shallow-water species, and acted as effective "geographic barriers" for pelagic, equatorial, Indo-Pacific stenothermal species during the Pleistocene. His arguments were based on the distribution patterns of calanoid (Calanoida; Calanidae) and pontellid (Calanoida; Pontellidae) copepods, which suggest that Wallacea may have been a long-term barrier, and played a significant vicariant role in speciation patterns of locally distributed species groups (Fleminger 1986).
The question we need to ask is what effect, if any, would low sea surface paleotemperatures, as suggested by Fleminger (1986), have had on the distribution and survival of shallow-water scleractinian corals in the much-reduced Pleistocene Wallacea? Even if we assume that sea surface paleotemperatures dropped seasonally to 20°C, the yearly average would still have been well within the lower tolerance levels for coral survival and reef development. However, low sea surface paleotemperatures as suggested by Fleminger (1986) are not supported by oxygen isotope data (518O) from deep -sea cores taken from two regions of Wallacea. Recent analysis of deep-sea cores taken from the northern regions of Wallacea (i.e., northern Molucca Sea) provided a wealth of information on the climatic conditions that existed during the last glacial maximum (Barmawidjaja et al. 1993). One of the most significant findings from the 5 O analysis was that surface seawater temperatures in the northern Molucca Sea during the last glacial maximum (i.e., 18 ka) were at most only 1°C cooler than at present. This suggests that glacial sea surface temperatures were basically within the optimal range for corals and coral reef development. 5 O analysis of deep-sea cores from the Banda Sea (i.e., southeast Wallacea) showed no evidence of significant reduction in surface seawater temperatures during the last glacial cycle (Ganssen et al. 1989). All recent evidence indicates that the average sea surface paleotemperatures in the archipelago most likely did not drop below 18°C, which is generally taken as the minimum for vigorous reef development.
Recent evidence against the presumed negative impacts of glacial temperatures on Indonesian corals and reef development can also be deduced indirectly from the occurrence of flourishing fringing reefs around many raised coral islands in the northern and southeastern regions of the archipelago (e.g., Sangihe Archipelago, Alor, Lembata, Adonara, Solor, etc.). It seems that coral reefs in these island chains are subjected to highly localized (scales 10's of m2) as well as highly transient (i.e., short duration with time scales of minutes to hours) tidally-induced upwelling events, which may lower sea surface temperatures to as low as 15°C during the Southeast Monsoon (K. Muller pers. comm.). These observations suggest that seasonal drops in sea surface temperatures may not necessarily inhibit reef development. With regards to the presumed increase in turbidity during the Pleistocene, it is worth noting that vigorous reef development in relatively turbid delta-front environments (e.g., northeast Kalimantan; Peleng Strait) suggests that sedimentation and/or turbidity were most likely of minor importance. Netherwood and Wight


Figure 6.5. Under certain environmental conditions, flourishing fringing reefs may develop adjacent to major river systems. A) This extensive fringing reef complex adjacent to a river is most likely maintained by strong coastal currents that keep fluvial sediments in suspension and facilitate rapid dilution of river runoff with coastal water masses. B) Coral community located approximately 200 m north (to the left) of the river delta shown in (A) above. The coral community is dominated by branching and massive Pontes spp., as well as robust Acropora spp. Heavy accumulation of siliciclastic sediments on the seaward slope of the fringing reef suggests that recent deforestation in the upper watershed has significantly increased sediment load in the coastal waters. Central Sulawesi, Peleng Strait. (1993) have recently described four sediment cores from the Tarakan Basin (northeast Kalimantan) that represent a carbonate interval (3466 - 3556 m) from the early Pliocene. Their interpretation of the cores was of four cycles of linear reef development on a shallow muddy shelf in a delta-front setting, a strikingly similar scenario that currently exists at the mouth of the Berau River. Flourishing fringing reefs have also been observed adjacent to rivers in Central Sulawesi (fig. 6.5).
Photos by Tomas and Anmarie Tomascik.
Umbgrove (1946c) supported Kuenen's general conclusions, by pointing out that the presence of 70%-100% of extant scleractinian species in the Pleistocene record does not support Daly's contention that the majority of corals were destroyed during the glacial maximum by either low temperature, or increased sedimentation (or turbidity) associated with erosion of the exposed foundations. It should be noted, however, that based primarily on data from Indonesian reefs, Daly (1948) conceded that coral reefs did not disappear in the core tropical regions. Kuenen (1933a) offered twenty solid arguments against the glacial-control theory and concluded that: "the formation of most atolls antedates the glacial period. They are essentially the products of reef growth combined with a sinking substratum". Thus, while accepting the fact that major eustatic sea-level changes have occurred, as was shown by Molengraaff (1916, 1917), Kuenen concluded that sea-level fluctuations had only a minimal effect on the already-existing atolls in the archipelago, whose formations predate the glacial period. He conceded, however, that Pleistocene sea-level lowstand may have had considerable influence on the uniformity of depth and the general morphology of barrier reef and atoll lagoons.
A few years later, Kuenen (1947) obtained new geologic evidence from two deep borings conducted on Maratua Island, which provided strong evidence for Darwin's subsidence theory. Based on the interpretation of the two deep cores from Maratua, and a review of the subsidence and glacial-control theories of atoll and barrier reef formation, Kuenen (1947) proposed "the theory of glacially controlled subsidence" (fig. 6.6).
Kuenen's (1947) hypothesis maintained that while pre-glacial reef development proceeds according to Darwin's subsidence theory, the present morphology of atolls and barrier reefs is due mainly to the effect of sea-level fluctuations. While appealing in concept, the theory did not receive wide acceptance mainly because, as pointed out by Scott and Rotondo (1983), it consists of two contrasting mechanisms (i.e., subsidence during Tertiary and modified glacial-control theory during the Pleistocene) to explain the same result. The major weakness of the glacially controlled subsidence theory, however, was the assumption that the Pleistocene foundations were cut down to low tide levels through chemical marine abrasion and denudation: "… thus producing hard rocky reef flats exactly at glacial sea level" (Kuenen 1947). In contrast to the glacial-control theory, where planation of subaerial foundations was primarily mechanical, the glacially controlled theory evoked subaerial limestone solution as the main process through which Pleistocene limestone foundations were truncated to the glacial sea level. Nonetheless, it seems indisputable that Kuenen's integrated hypothesis for the formation of barrier reefs and atolls in the Indonesian Archipelago explained many geomorphological features (e.g., steep outer reef slopes, presence of barriers and atolls on submarine ridges, uniformity of lagoonal depths, etc.; see Kuenen 1947), which neither the subsidence nor glacial-control theories could explain alone. In his 1947 review Kuenen concluded that:

Figure 6.6. The theory of glacially controlled subsidence. A) Pre-glacial reef development: a) barrier reef; b) fringing reef; c) lagoon and lagoonal sediments; d) fore-reef talus. B) Glacial maximum and sea-level lowstand: a) living coral at the edge of abraded barrier reef; b) remnants of a fringing reef; c) aggraded lagoon; d) fore-reef talus. C) Interglacial: a) new barrier reef; b) new fringing reef; c) lagoon and new lagoonal deposits; d) fore-reef talus; e) lagoonal reef.
Modified from Kuenen 1947.
It must be left to future investigation to determine in how far the chemical low-level abrasion was able to destruct former reefs and to show whether these were entirely planed off or only moderately destructed.—KUENEN 1947
The Antecedent-Platform Theory. As was recognized by Hopley (1983), implicit in Kuenen's glacially controlled subsidence thesis was the role of chemically denuded Pleistocene foundations on which coral reefs subsequently took hold. However, even though Kuenen accepted the importance of antecedent platforms in the formation of recent barrier reefs and atolls, he disagreed that the antecedent-platform theory of Hoffmeister and Ladd (1944) can be considered as an alternative to Darwin's subsidence theory. As Guilcher (1988) pointed out, the antecedent-platform theory evolved from the works of such notables as Semper (1863), Murray (1889), Wharton (1897), Gardiner (1898,1904), Wood-Jones (1910) and Vaughan (1919, 1923). Briefly stated, the antecedent-platform theory proposes that barrier reefs and atolls can develop on any suitable pre-existing platform provided it is located within the euphotic zone where environmental (biotic and abiotic) conditions permit coral recruitment and growth. More to the point, Hoffmeister and Ladd (1944) disagreed with the genetic implications of Darwin's theory, and maintained that both barrier reefs and atolls may develop simply by vertical growth of corals around the seaward edges of antecedent platforms, thus eliminating the need for fringing reefs. Furthermore, they believed that once corals become established, vertical reef growth occurs independent of sea-level changes. Hoffmeister and Ladd (1944) obtained evidence for the antecedent-platform theory from their work in the Fiji Islands. They suggested that most volcanic islands will be rapidly abraded by wave action to a few dozen metres below sea level once volcanic activity ceases. Aggradation of the planed-off foundation proceeds through deposition of fine limestone sediments (e.g., foraminifera, molluscs, algae, etc.) to a depth where corals are able to survive and reef development can begin around the edges of the platform. The upward growth of coral will subsequently result in the formation of barrier reefs and atolls. It is important to point out that Hoffmeister and Ladd (1944) did not rule out subsequent growth of the barrier reefs and atolls through further subsidence of the platform.
The theory also assumed that most oceanic volcanic islands were at the mercy of the sea, since erosion was evoked as the major process through which these foundations became truncated many metres below sea level. Our recent observation of Banda Api volcano, however, clearly demonstrates that, depending on the nature of the volcanic activity, fringing reefs may develop around active volcanoes, thus providing substantial protection against wave abrasion, thus considerably slowing or retarding erosion of the island. A number of volcanic islands in the Banda Sea (e.g., Damar, Gunung Api, Serua, etc.) are surrounded by well developed fringing reefs that offer considerable protection to their coastlines. These observations illustrate that planation of volcanic islands prior to reef development is not necessarily true, as has been demonstrated by the Mururoa borings. It is highly unlikely that the antecedent-platform theory, as viewed by Hoffmeister and Ladd (1944), applies to Indonesian barrier reefs and atolls.
The Karstic-Saucer Theory. Daly's (1915) seminal publication of glacial-control theory of coral reef development, while heavily criticized, brought to light the effects of Quaternary climatic and sea-level changes, a concept that became a foundation of the karstic-saucer theory. While the 1951 Enewetak drilling program proved the main thesis of Darwin's subsidence theory, the evidence of major Quaternary sea-level fluctuations and its implication for Quaternary reef development could not be ignored. One of the major weaknesses of Daly's theory, as well as of the earlier antecedent-platform theories, was the assumption that coral reefs, and other foundations of present-day barrier reefs and atolls, were totally or at least partially truncated by mechanical wave abrasion to the lowest Quaternary sea levels. As Hopley (1982) pointed out: "... no link was made between the exposure of the limestone structures of reefs during low sea levels, and the development of solution-dominated karst landforms on this limestone". The concept that karst topography-a common feature on many islands throughout the Indo-Pacific (e.g., Okinawa, Palau, Kakaban, Maratua) - of antecedent-platforms was related to the general morphology of atolls was first introduced by Yabe (1942) and Asano (1942). However, considering the timing of their publications, their views went totally unnoticed by the international scientific coral reef community until an English translation of Japanese contributions to coral reef science was made by Burke (1951). In the interim, however, parallel ideas were advanced by Hoffmeister and Ladd (1945) to explain the origin of emerged coralline limestones in the western Pacific. Supported by their laboratory studies in which they exposed limestone blocks to dilute acid rain, they concluded that the raised coral islands were not uplifted atolls, as was originally believed, but rather that the atoll-like surface features were due to solution subsequent to subaerial exposure (Hoffmeister and Ladd 1945). Working on the coral reefs in the Indonesian Archipelago, both Umbgrove (1947) and Kuenen (1947) considered the effects of chemical solution, and its effect on limestone structures, but both rejected the idea that the general features of Indonesian atolls were related to this process. Nonetheless, the chemical solution effect of seawater on emerged intertidal reef and limestone structures was, however, considered by both as an important process in the formation of coastal features. Kuenen (1947) was of the opinion that coastal features, such as mushroom-shaped rocks and deep notches that undercut even well protected, emerged reefal limestones (fig. 6.7) were a result of chemical abrasion. However, considering the fact that the mushroom-shaped rocks were undercut just about evenly from all sides, Kuenen (1947) did not consider mechanical attack to be an important process in their formation. Addressing the problem that tropical reefal waters are generally oversaturated with respect to CaCO3, Kuenen (1947) suggested that: "… it is very well possible, that after rain, or during night or storm, a temporary undersaturation is developed". Indeed, it is now generally accepted that diurnal variations in pH do occur in microenvironments such as algal mats, interstitial spaces, or in the films of surface water left on intertidal reefal limestone by receding tides (Hopley 1982).
Trudgill (1976) demonstrated that, while chemical solution of reefal limestone does occur, it is generally insignificant when compared to bioerosion. In contrast to Kuenen's (1947) conclusion that chemical solution was the main process behind the formation of notched limestone cliffs, Umbgrove (1947) suggested that the notches and mushroom-like formations originate as a result of: "… the sawing action of the sea and boring by the organisms…". The role of boring organisms was later supported by the work of Hodgkin (1970), who demonstrated that the notches (e.g., see fig. 6.7) are primarily bioerosional features. Our direct observations of mushroom-shaped formations and intertidal notches at Kakaban and Maratua Islands (East Kalimantan), Banggai Island (Central Sulawesi), the Banda Islands (Moluccas) as well as Palau (Caroline Islands), confirm Hodgkin's (1970) conclusions that endolithic algae, chitons, echinoderms, boring sponges (predominantly Cliona spp.), boring bivalve molluscs (e.g., Lithophaga) and a host of other boring organisms are the primary agents responsible for these characteristic features of coral coastlines. Kuenen (1947) commented that: "…chemical attack on elevated limestones by weathering cannot be doubted…", and suggested that production of humic acids by the tropical vegetation covering exposed reef rock will greatly intensify the dissolution process. However, the absence of emerged outcrops of interglacial reefs from present-day reefs, in spite of the fact that interglacial sea levels were above present, led Kuenen (1947) to conclude that Pleistocene reefs were mostly abraded during low sea-level periods.

Figure 6.7. Many emerged limestone cliffs in the Indonesian Archipelago have a characteristic intertidal notch and visor which is caused primarily by the combined action of physical (abrasion), chemical (solution) and biological (bioerosion- physical and chemical) processes. Recent evidence indicates that bioerosion, rather than chemical solution or wave action, is the primary mechanism of formation.
Photo by Tomas and Anmarie Tomascik, southwest coast of Banggai Island, Central Sulawesi.
Kuenen's (1947) glacially controlled subsidence theory was an attempt to reconcile two indisputable facts, namely the evidence of general subsidence and the Pleistocene sea-level fluctuations. The integration of subsidence and sea-level fluctuations was finally placed into a proper framework by Tayama (1952), who suggested that Quaternary reefal build-ups may have occurred mostly on previously emerged and karstified older reefs. These ideas were developed further by MacNeil (1954), who proposed that atolls inherited their annular shape primarily from their karstified antecedent-platforms, as well as to reef growth during periods of submergence. Note that MacNeil (1954) clearly recognized that a single theory to explain barrier reefs and atolls may not be realistic, and stated that: "There is a general feeling among investigators at the present that atolls are not always formed the same way, and therefore no one theory can explain atolls". Interestingly enough, MacNeil (1954) used the raised atolls of Kakaban and Maratua as well as the submerged Muaras Atoll as evidence that the theory is also applicable to shelf reefs. However, Kuenen (1947) provides a strong argument suggesting that both Kakaban and Maratua were formed during slow subsidence of their foundation, which was subsequently uplifted by tectonic forces.
The karstic-saucer theory was greatly expanded upon by E.G. Purdy, who obtained additional support for the general aspects of the theory from both laboratory solution studies (i.e., limestone blocks exposed to acid rains) (Purdy 1974a), as well as from his work on the Belize Shelf (Purdy 1974b). The karst-induced antecedent-platform theory is applicable for the formation of barrier reefs and the enclosed lagoonal patch reefs (fig. 6.8), as well as for the formation of atolls (fig. 6.9).
The theory's basic premise is that the characteristic annular (i.e., saucer-like) shapes of atolls, as well as the general surface features of barrier reefs, are derived from a generally flat, carbonate antecedent platform, which, while subaerially exposed during relative sea-level drops, was karstified by rainfall and percolating water (Guilcher 1988). The karstification of the exposed limestone produced a characteristic saucer-like morphology, with a central lagoon with numerous depressions surrounded by a slightly elevated outer rim breached by gullies (i.e., deep passes), which presumably acted as drainage streams during subaerial exposure. The karstification also produced features such as inter-reef channels, drowned dolines and blue holes (figs. 6.10, 6.11). Blue holes are common in many reef types (e.g., fringing reefs, barrier reefs and atolls) throughout the archipelago, and are a key feature in Purdy's (1974a) defense of the karstic-saucer theory. Since blue holes have thus far not been investigated in Indonesia, their origin remains generally unknown. Hopley (1982), however, suggested that, with regards to blue holes on the Great Barrier Reef, their origin is most likely much older than the last interglacial.
Following the karstification of exposed limestone foundations during sea-level lowstands, reef development occurred during transgressions over the karstified limestone antecedents. The theory proposed that reef growth on the annular karstified rim resulted in the morphology characteristic of atolls and numerous lagoonal shelf reefs. The upgrowth of reefs may occur as a result of eustatic sea-level rise, subsidence of the foundation, or both; thus it is not in contradiction to Darwin's subsidence. The major thesis of the karst-induced antecedent-platform theory lies with the formation of the atoll rim, and it is here that it is in contradiction with Darwin's hypothesis, which maintains that the rim is a result of vertical reef growth on a subsiding foundation. The best evidence for the karst-induced antecedent-platform theory comes from Purdy's (1974a) work on the barrier reef off Belize (see Guilcher 1988). By the early 1980s the theory became so well established that Scott and Rotondo (1983) concluded: "… it is clear that the present atoll morphology is fundamentally karst induced, rather than growth induced". It is noteworthy to point out, however, that Hopley (1982) in his exhaustive review of the antecedent-karst theory stated that, in general, it cannot be sustained on the Great Barrier Reef. Hopley provides strong arguments in support of his conclusions, namely: 1) the unsuitability of Quaternary reefal limestone to form karst topography; and 2) low rates of karstification of young Quaternary reefal limestones. In his concluding remarks on the foundation of modern reefs, Hopley (1982), referring to the importance of solution during subaerial exposure, stated:

Figure 6.8. Generalized interpretation of Purdy's (1974a) karst-induced antecedent-platform theory as applied to the formation of barrier reefs.
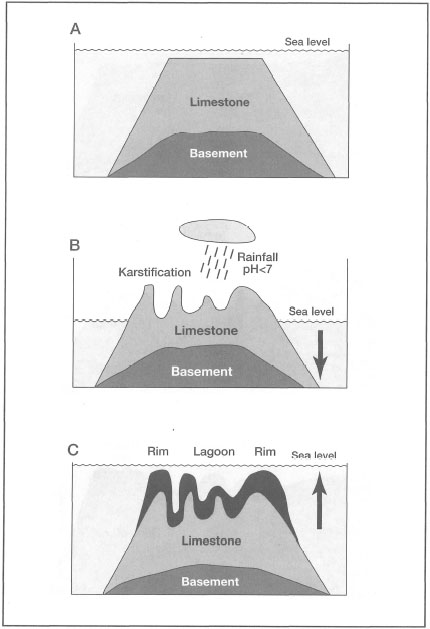
Figure 6.9. Generalized interpretation of Purdy's (1974a) karst-induced antecedent-platform theory as applied to the formation of atolls.

Figure 6.10. An aerial photograph of a narrow fringing reef along the south coastline of Tomini Gulf, Central Sulawesi. Four blue holes are clearly visible, lower centre. Their presence on a relatively narrow fringing reef suggests that they are solution features.
Photo by Tomas and Anmarie Tomascik.

Figure 6.11. An aerial photograph of an unusual reef complex along the south coastline of Tomini Gulf, Central Sulawesi. Numerous blue holes associated with the fringing reef are visible. The small barrier reefs are of unknown origin. Depth of lagoons and blue holes is unknown.
Photo by Tomas and Anmarie Tomascik.
Relief features may have been enhanced over several glacial-interglacial cycles, as subaerial processes may be guided by contrasts in original reef facies. But more influential in providing the relief of both the pre-Holocene foundation and the Holocene morphology is the pattern of growth during submergence that will favor the colonization of prominences at the expense of depressions.
The strongest evidence against the karst-induced antecedent-platform theory derives from the results of numerous borings at Mururoa Atoll, which became available during the 5th International Coral Reef Congress in Tahiti (Buigues 1985). The deep cores revealed that while the lagoon is primarily made of fine muddy sediments, the outer rim of the atoll consists of built-up reef and coarse slope debris (Buigues 1985). The most significant fact is that these differences have persisted since well before the Holocene (Guilcher 1988). However, on a more cautious note, Guilcher (1988) concluded that: "Beside the general fact of subsidence combined with variations in sea-level, and tectonic uplift in a number of cases, it appears wise to accept the possibility of more than one explanation of reef rim formation until many more borings have been made".