Chapter Twenty-One
INTRODUCTION
The Indonesian Archipelago is one of the richest marine biodiversity areas in the world. While the total biodiversity of any particular area is a function of a complex set of physical, chemical and biological interactions, as well as geological history, it will be useful here to discuss biodiversity at two levels, namely at the ecosystem level and the species level. Genetic biodiversity is the least understood level and is beyond the scope of this book.
The exceptional biodiversity of marine flora and fauna in the Indonesian Archipelago is a result of its geographic location and geologic history. The vast archipelago can be viewed as a mixing pot of a number of biogeographic provinces. In the previous chapters we examined the physical and chemical factors that support this profusion and diversity of marine life, and now we focus our attention on the distribution and abundance of marine life that abounds in the archipelagic waters. Because of the nature of this book our main focus will be on a small group of marine organisms that have significant national and global conservation value. There is some literature dating back to the 16th century that describes the fascinating nature of marine life in this part of the world. With the relatively recent advent of scuba (self-contained-underwater-breathing-apparatus), the volume of research articles has grown considerably, many of which are in the so-called 'grey literature'. However, the marine biodiversity of this complex archipelago is still poorly known and documented. Large marine groups such as the pelagic jellyfish (Mass 1903) and ascidians (Sluiter 1903) remain poorly known in Indonesian waters. Major revisions of many marine taxa since the Siboga and Snellius-II Expeditions make the expedition reports very difficult to use by non-specialists. Most of the present knowledge of marine life in Indonesian waters comes from the shallow-water environments (i.e., the shelf seas) of less than 200 metres depth. This is primarily due to the fact that the Indonesian coastal zone, one of the largest in the world, supports a high diversity of productive ecosystems (e.g., wetlands, estuaries, beaches, mangroves, coral reefs, seagrass and algal beds, etc.), which, in turn, support a wealth of marine resources.
The terrestrial fauna and flora of the Indonesian Archipelago are characterized by high endemicity. For example, about 50% of Indonesian primates are endemic. Java, alone, accounts for about 325 endemic plant species and 31 endemic birds, and Borneo boasts about 60 endemic plant genera (KLH 1992). In comparison, the endemism in the marine environment is much less pronounced. However, this view may change, since new genera and species are being described each year from remote parts of the archipelago (e.g., Ng and Tomascik 1994; Wallace 1994, in press; Kott 1995; Massin and Tomascik 1996). The marine species list will grow considerably as newly trained taxonomists tackle large volumes of material collected during the various expeditions in the past. With regards to corals alone, G. Wallace has recently described eight new endemic Acropora species from various parts of Indonesia. Some of these new endemics are specific to certain parts of the archipelago. For example, two endemics are restricted to the Gulf of Tomini (e.g., Togian Islands) and one to the Sulawesi Sea (e.g., Berau Islands), while others have wider distributions throughout the archipelago (Wallace, in press). One new endemic, a large table species, is found throughout the archipelago, yet it has not been found in the Philippines or anywhere else outside Indonesian waters (Wallace, in press). The biogeographical distribution of corals is also changing rapidly as new records are being made. Even in intensively studied areas, such as Kepulauan Seribu and the Spermonde Archipelago, new coral records are still being recorded (see box. 7.2 chapter 7).
BlOGEOGRAPHIC CLASSIFICATION
The coastal and marine environments of the world were divided by Ekman (1953) into the Indo-west Pacific and the Atlanto-East Pacific components, the latter being further divided into Tropical and Subtropical America and Tropical and Subtropical Africa. Forest and Guinot (1962) separated the Tropical and Subtropical America into Tropical and Subtropical West Atlantic and Tropical and Subtropical East Pacific. The Indo-west Pacific region is vast, comprising tens of thousands of islands, many of which are coral islands. The region is centred in the Malay Archipelago and extends west across the northern part of the Indian Ocean to the eastern coast of tropical Africa and the Red Sea. From its western boundary it extends eastward to the Tuamotu Archipelago in the South Pacific, and northward to southern Japan and southward to the southern tip of Australia.
One of the main marine biodiversity centres of the Indo-west Pacific region is the Malayan Archipelago. Ekman (1953) and Briggs (1974,1987) consider this area as the faunistic centre from which the other regions of the Indo-west Pacific recruited their faunas. This hypothesis is based on the presence of advanced species in the Malay Archipelago and more primitive ones occupying peripheral habitats (Briggs 1987). However, the area could not be considered as a distinct biogeographic province, mainly due to the lack of endemism (Briggs 1987). Briggs (1974, 1987) placed the Malayan Archipelago (i.e., East Indies triangle), with its widely distributed marine fauna, at the heart of the Indo-Polynesian Province of the Indo-west Pacific biogeographic region.
Briggs's (1974) modification of Ekman's (1953) system now stands as the primary marine biogeography classification on a world level (Hayden et al. 1984). Realms are considered as major subdivisions of the world, based mainly on the distribution of species. Three realms are recognized, namely, the continental shelf realm, the pelagic realm (epipelagic, mesopelagic and bathypelagic zones) and the deep benthic realm (abyssal plains, hadal zone - deep trenches). Each realm in turn consists of one or more regions, which are usually large and specific areas within each realm. Provinces refer to areas within a region, and their boundaries are generally delineated by the distribution of endemics or characteristic biotic associations (Hayden et al. 1984).
Even though biogeography is a well-established discipline, there are surprisingly few biogeographical classifications of marine areas. Hayden et al. (1984) noted that the difficulties in preparing a marine biogeographic classification are substantial, and that the physico-chemical domains of the oceans, such as temperature, salinity, and surface currents, are generally agreed to be the basis of classification systems. The World National Congress in Bali, in 1982, identified specific needs, one of which was to "develop an appropriate marine biogeographical classification scheme on global, regional and national levels as a basis for ensuring adequate representation of different marine ecosystems in a wide range of protected areas."
Kelleher et al. (1995) produced maps of 18 biogeographical marine regions as part of an international effort to conserve marine biodiversity and achieve sustainable use of the marine environment. Indonesia is placed in Marine Region 13 (i.e., East Asian Seas). However, they also noted that the marine environment has proved to be much more difficult to categorize according to biogeographic regions, and that there is no general consensus on any one system (Kelleher et al. 1995). The coastal zone is considered to be of highest priority.
Bleakley and Wells (1995) included the following countries in the East Asian Seas Marine Region No. 13: Indonesia, Malaysia, the Philippines, Singapore, Brunei Darussalam, Thailand, Cambodia and Vietnam. The East Asian Seas Marine Region comprises the Andaman Sea, Straits of Malacca, Straits of Singapore, South China Sea, Java Sea, Flores Sea, Banda Sea, Arafura Sea, Timor Sea, Celebes Sea, Sulu Sea, and the Philippine Sea (and all other ASEAN intra-archipelagic seas). The East Asian Seas Marine Region includes shallow continental shelves, deep-sea basins, troughs, deep-sea trenches, continental slopes and volcanic islands. The numerous large and small islands divide the marine waters of the region into different seas connected by many channels, passages and straits. Classification of biogeographic subdivisions for Indonesia was proposed by Hayden et al. (1984), however, its value is still to be determined. Bleakley and Wells (1995) have recently subdivided the Indonesian Archipelago into seven first-order biogeographic subdivisions and 11 second-order biogeographic subdivisions. While these classifications may be useful in conservation and management of the diverse marine and coastal resources, they have very little to do with biogeography of Indonesian marine flora and fauna.
Coastal Ecosystems
Soegiarto and Polunin (1981) provided the first detailed review of the Indonesian marine environment. Their work was followed by Salm and Halim (1984), who proposed a preliminary classification for a number of marine and coastal habitats throughout the archipelago. Their marine atlas is frequently cited in conservation and coastal zone management literature. The status of Indonesian marine and coastal resources was subsequently reviewed by Polunin (1983) and Knox and Miyabara (1984). In their review of Indonesian coastal zone management development, Burbridge and Maragos (1985) identified 11 key marine and coastal ecosystems that were categorized as critical for future national development; they are summarized in table 21.1. While considerable progress has been made in acquiring new information on coral reefs, mangroves and seagrass beds, very little information has been added on beaches, sand dunes, rocky shores and many other marine ecosystems. Coral reefs, mangroves and seagrass beds and associated habitats have been discussed in earlier chapters and will not be included here. The following brief discussion is based primarily on the work of Soegiarto and Polunin (1981), and provides only a cursory treatment of three ecosystems where new research is urgently needed.
Table 21.1. Summary of critical marine and coastal ecosystems in Indonesia.


Beaches. As the sharp boundary between the land and the sea, beach and dune ecosystems show a distinct ecological zonation of communities as well as productivity (Soegiarto and Polunin 1981). The beach is often regarded as an amalgam of sub-, inter- and supra-tidal zones, embracing nearshore, surf and swash zones between the LW breaker line and the HW run-up limit (Carter 1988). These systems are important components of the Indonesian coastal zone. Beaches show considerable variation in their profiles and can change markedly from season to season. In general, there is a direct relationship between beach zonation and exposure. Dunes are apparently infrequent in Indonesia, since they form mainly on dissipative coasts with strong onshore winds. These conditions exist along the south coast of Java as well as along the north coast of Irian Jaya. However, no information is available on their status.
In the more stable supra-tidal zone strand plants dominate, while in areas where the beach is accreting, the outer fringe of vegetation most commonly comprises a pes-caprae community, named after the characteristic species Ipomoea pescaprae (Soegiarto and Polunin 1981). The plants in this community are low sand-binding herbs, grasses and sedges. J. pes-caprae often predominates in the seaward pioneering fringe, while the grasses and sedges such as species of Cyperus, Fimbristylis and Ischaemum occur landward (Soegiarto and Polunin 1981). As beach pioneers, the plants are commonly deep-rooting, are tolerant of salt (although dependent on non-saline ground water), wind and high soil temperatures, and produce floating seeds (Soegiarto and Polunin 1981). However, beach vegetation is usually species-poor and often communities are dominated by one or a few species. On many coasts almost pure stands of Casuarina equisetifolia are common, and these can often be succeeded by a full Barringtonia community (Soegiarto and Polunin 1981).
Stable soils behind the beach (or on rocky soils where there is no beach accumulation) are generally colonized by trees and shrubs which form the Barringtonia community, one characteristic species being Barringtonia asiatica (Soegiarto and Polunin 1981). In undisturbed areas, the tree canopy can be dense, and there may be some understory vegetation present. In clearings and open spaces, ferns, grasses, gingers and herbs dominate (Soegiarto and Polunin 1981). The community comprises rather few species, but most of these are restricted to this habitat, although many are widely distributed throughout the tropics (Soegiarto and Polunin 1981).
Exposed beaches represent a particularly hostile environment (due to physical instability, and wide variations in temperature, salinity and humidity), however, a number of animals have adapted to these conditions (Soegiarto and Polunin 1981). The main factor determining the populations within the beach is grain size (Carter 1988). There is a strong environmental gradient from the exposed seaward part of the beach to the sheltered, landward part of the beach. The energy gradient is reflected in the distribution of sediment grain size. Sediments in the high-energy (i.e., exposed) zone consist predominantly of large-size grains (up to boulder size) which grade into gravel, sand, silt and mud as one moves from the high-energy to low-energy conditions.
Beach organisms are frequently divided into three sediment-size related groups: 1) attached microfauna; 2) interstitial meiofauna; and 3) burrowing or loosely attached macrofauna (Carter 1988). Very little is, however, known about Indonesian beach fauna. The beach and nearshore surface is inhabited by detrivorous and predatory crustaceans, molluscs, echinoids, fish and birds (Carter 1988). Many inhabitants of the beach follow the tidal cycle by moving up and down the beach. Beaches that are fronted by seagrass beds often support large populations of scavengers which feed on the decaying organic material. The large animals occurring on the beach generally show a well-developed zonation; ghost-crabs of the genus Ocypode and amphipod crustaceans (Talitridae) are generally characteristic of the upper zones, the middle zones are usually colonized by the mollusc Donax and isopod crustaceans such as species of Exirolanus, and the lower zone is characterized by the gastropod Ombonium, hippid crabs and the sea urchin Echinodiscus (Soegiarto and Polunin 1981). Beaches are also important nesting habitats for sea turtles, and megapode birds in the eastern parts of the archipelago (e.g., Banggai Islands) (Soegiarto and Polunin 1981).
The nearshore waters support rich populations of phytoplankton and zooplankton, which derive, great benefit from turbulence generated by the continual wave action, and longshore currents. Diatoms are the most abundant indigenous phytoplankton group (e.g., Chaetoceros). Diatoms thrive particularly in the surf zone of high-energy beaches (e.g., south coast of Java and Bali) where nutrients are available from land runoff and strong coastal upwelling. Surf-zone diatoms have the ability to attach themselves to bubbles, which allows them to remain at the surface in maximum light while taking advantage of longshore currents and prevailing surface flows (Carter 1988). Not much is known about the nearshore productivity of beach ecosystems in Indonesia, but Soegiarto and Polunin (1981) reported that in west Bali, the beach seine fishery (Stolephorus, Sardinella and Leiognathus) is an important coastal activity. Furthermore, nearshore beach areas are important habitats for the commercially important milkfish (Chanos chanos) fry that aggregate in these areas for a few days during part of their life cycle before returning to deeper offshore waters (Soegiarto and Polunin 1981).
Rocky Shores. Rocky shores occur along the coastlines of many Indonesian islands. Long stretches of coastline along the west coast of Sumatra are lined by rocky beaches, as are numerous volcanic and non-volcanic islands in the Lesser Sunda Islands and the Moluccas. Whitten et al. (1987) mention rocky shores consisting primarily of limestone along the western shore of Aceh, volcanic rocky shores south of Padang (West Sumatra) as well as rocky shores consisting of Tertiary sandstone in Belitung and many other high islands. Many islands whose shores are exposed to strong wave action and currents have steep cliffs along their coastlines with little or no beach formation.
The zonation of organisms on rocky shores usually follows the typical pattern with three major zones, namely sub-, mid-, and supra-littoral. The organisms that inhabit rocky shores are either sessile or mobile. Each zone is characterized by a key group of organisms such as the littorinids (e.g., Littorina spp. and Nodolittorina spp.), gastropods (e.g., Trochus maculatus, Eucheus atratus, Nerritapolita, N. plicata),chitons (Acanthopleura spp.), barnacles (pedunculate and sessile; e.g., Tetraclita porosa, T. costat, T. coerulescens, T. vitiata), and algae. Space is a key limiting factor in the rocky intertidal, and competition for this resource among sessile organisms, such as barnacles and oysters, can be intense. While the floral and faunal diversity of many rocky shores is higher than that of sandy beaches, it is nonetheless not particularly rich, even though it may contain representatives from a wide range of phyla (Whitten et al. 1988). The habitat found between the high and low water (i.e., intertidal) is a steep ecocline, where sharp changes in environmental conditions occur across the shore (Morton 1990). High temperatures and desiccation are the key environmental factors that limit the number of fauna and flora. The rock pools which are subject to extreme heating during the day contain animals which live mostly in crevices, such as shrimps (alpheids), ophiuroids and snails. Larger intertidal pools, however, support a host of small fish such as gobies and blennies.
Rocky shores are influenced by both tides and waves. In many areas (e.g., south coast of Java), waves actually have greater amplitude than tides, continually wetting the intertidal and supratidal areas. However, the actual vertical range of this particular habitat is shorter in the tropics than in the temperate zones (Morton 1990). In Indonesia, study of rocky shore zonation has been greatly neglected, and very little is actually known about their ecology (Whitten et al. 1987; Whitten et al. 1988). Past studies were concerned mainly with species collections and not ecological interactions. The south coast of Java, with many easily accessible rocky shores, is an ideal natural laboratory to initiate research into rocky shore ecology.
Intertidal Mud Flats. Mud flats are categorized as tidal flats (i.e., flat soft-bottom habitats between the low and high water). Intertidal mud flats are most frequently built in estuarine basins, and are depositional features composed of loose, coarse and fine sand and silt. They are common in large estuaries where they often divide, or braid, the original channel. Whether intertidal mud flats become vegetated depends primarily on substrate composition and tidal action. Under natural conditions, intertidal mud flats are an important habitat for a variety of organisms that feed on organic matter brought in by the tides and river runoff, or on autochthonous production of organic detritus.
These soft-bottom habitats are inhabited by a variety of marine invertebrates, algae, seagrasses and higher plants (e.g., mangroves). Benthic organisms inhabiting intertidal mud flats can be divided into three main groups, namely the sessile epibenthos (e.g., barnacles, seaweeds, crinoids, seagrasses, oysters, clams, anemones, sponges, etc.), mobile epibenthos (e.g., gastropods, starfish, and nudibranchs) and the infauna (e.g., mussels, worms, crabs, etc.). Gastropods are commonly the most abundant group of macroinvertebrates on many intertidal mud flats. Other invertebrates present in high numbers are crustaceans, polychaetes, sipunculides, bivalves, coelenterates, and echinoderms. Several gastropods and bivalves (e.g., Pinna bicolor, Anadara granosa, Paphia luzonica and Laternula truncata) are commonly found burrowing in sandy or muddy substrates. Burrowing deep into the sediments allows burrowing organisms to avoid heat exposure during low tides when significant heating of substrate occurs. Sea cucumbers (e.g., Holothuria spp.), and starfish, such as Linckia sp. and Ophiotrichoides sp., are also abundant components of many sand flats or mud flats. Crabs, such as Uca spp., are among the most common motile fauna of intertidal mud flats and sand flats.
Because of their high productivity, intertidal mud flats are important feeding habitats for thousands of resident and migratory waterbirds. However, with rapid coastal development many of these unique habitats are being degraded at the expense of many rare bird species. Most of our current knowledge on intertidal mud flats and sand flats relates to species compositions, with only scant information on ecological interactions. Research is needed, especially since intertidal mud flats are highly susceptible to land-based pollution.
MARINE FLORA AND FAUNA
The Indonesian Archipelago was visited by numerous expeditions which collected an impressive volume of information, most of it stored in specimen bottles. Only a few Indonesian scientists have been able to study these collections. The field of marine taxonomy has been generally neglected in Indonesia, as other, more pressing, needs required attention. However, with new emphasis on sustainable development, and on the management and conservation of marine biodiversity, the need for taxonomists will continue to increase. Currently, there are fewer than five Indonesian taxonomists working with scleractinians, even though information on Indonesian species is essential to understand large-scale regional and global biogeographical distribution patterns. While a large number of surveys are being done throughout the archipelago, they are often cursory in nature. Indonesian coral reefs probably have the most diverse fish fauna in the world, yet few species lists are available. Maumere Bay alone is home to over 1700 species offish (G. Allen, pers. comm.), however, no reports are available. The large Indonesian Octocorallia and Porifera collections stored at the National Museum of Natural History in Leiden and Amsterdam Zoological Museum, respectively, have yet to be studied in detail by Indonesian scientists. It is, therefore, not surprising that our current knowledge of marine biodiversity is poor, and clearly insufficient for management purposes.

Figure 21.1. Large barrel sponges such as this Petrosia testudinaria are important components of coral reef ecosystems. Berau Islands, East Kalimantan.
Photo by Tomas and Anmarie Tomascik.

Figure 21.2. Triphyllozoon sp., the Bryozoa, commonly called the 'sea mosses', are a fascinating group of marine organisms. Bryozoan colonies consist of thou sands of microscopic zooids (individuals) that form visible colonies reinforced by calcium carbonate. They are an abundant group of benthic coral reef organisms. Damar Island, Banda Sea.
Photo by Tomas and Anmarie Tomascik.
Indonesian coral reefs are, without a doubt, the highlight of Indonesian marine biodiversity. In addition to the 450 or so reef-building coral species found in Indonesian waters, there are a multitude of other reef invertebrates such as sponges (fig. 21.1), benthic foraminiferans, bryozoans (fig. 21.2), polychaete worms (fig. 21.3), crustaceans, molluscs, holothurians and echinoderms, all of which are important biological components of Indonesian coral reef ecosystems. These organisms, however, are not considered as reef-builders (i.e., constructional or hermatypic), even though their skeletal remains may contribute significantly to reefal sediments.

Figure 21.3. Serpulid worms (Polychaeta, Serpulidae) are an abundant group of reef organisms. Filograna sp. form delicate branched colonies. They can be locally very abundant, and contribute to the production of reefal sediments.
Photo by Tomas and Anmarie Tomascik.
Of the hundreds of genera of reef invertebrates that produce calcareous shells, tests, spicules and various other kinds of skeletal material, well over 100 genera are symbiotic with different endosymbionts (e.g., cyanophytes, dinoflagellates, dinomastigotes, chlorophytes, etc.). The symbiotic relationship that has evolved in these various groups of reef organisms is functionally very similar to the scleractinian-zooxanthellae symbiosis. However, with the exception of foraminifera, radiolaria and the tridacnids, not much is known about the functional role of symbiosis in the other major groups of marine organisms.
Among the most unique symbiotic relationships on the reefs is that of the ascidian Didemnum molle (Didemnidae) and its Prochloron photoendosymbiont. The zooids of all Didemnidae are very small, and the colonies are frequently mistaken for sponges by divers (colour plate 21.1). Kott (1980, 1982) made a revision of the Prochloron-containmg didemnids. Prochloron is an unusual group of oxygenic photosynthetic prokaryotes containing chlorophyll-a and -b in paired thylakoids, but they lack phycobilins which are present in the cyanobacteria. Thus, while the cellular organization of Prochloron is prokaryotic, they differ from cyanobacteria in their photosynthetic pigments (Delaney 1990). Prochloron has been placed in the Division Prochlorophyta. Recently, a new type of prochlorophyte was discovered, which differs from Prochloron by being both filamentous and planktonic; also, it occurs in reefal waters (Burger-Wiersma et al. 1986). The larvae of Didemnum mollehave special "lunch boxes" (Steene 1990) by which they carry Prochloron with them (colour plate 21.1B). Another interesting feature about Didemnum molle is that, unlike many other ascidians, the colonies are motile (fig. 21.5). Limited locomotion is achieved by sending out thin, white fibers that attach to the substrate some distance from the colony and subsequently act as anchor lines along which the colony pulls itself to a new location (fig. 21.5). Locomotion of Diplosoma virenswas observed under laboratory conditions and measured at rates of 4.7 ± 0.53 mm. 12hr (Birkeland et al. 1981). It is believed that the motility allows the colonies to position themselves in light conditions favourable to their Prochloron symbionts.
Box 21.1. The algae.
W.F.Prud'homme van Reine, Research Institute Rijksherbarium Hortus Botanicus, Leiden University, Leiden, the Netherlands.
Marine Algae
Introduction
Algae are oxygen-producing, photosynthetic organisms with unprotected reproductive structures. They are often simply constructed, ranging from single-celled forms to cell colonies, filaments, or parenchymatous thalli. Algae, as such, do not form a specific taxonomic group, but include all plant-like prokaryotic and eukaryotic form organisms that are neither mosses nor vascular plants. Seagrasses, however, belong to the vascular plants since they possess vascular tissue.
Photosynthetic Pigments
Algae, being photosynthetic organisms, need light to survive. Special pigments are used by the different groups of algae to harness light energy in the process of photosynthesis. Most algal groups are named after the colour of their dominant pigments. In all algae, the main pigment is chlorophyll-a, which is green in colour. However, most algae with chlorophyll-a also contain other photosynthetic pigments, such as the red, orange, yellow or brownish fat-soluble carotenoids. These accessory pigments mask the green chlorophyll-a, which gives many different algal groups their specific colouration.
In the cyanobacteria, additional pigments are mainly water-soluble phycobilins, viz. the bluish phycocyanins and the red phycoerythrins. In the red algae the phycoerythrins dominate, while in the brown algae the presence of the carotenoid fucoxanthin results in dark brown thalli. In the green algae the colour of the chlorophyll-a is not masked.
Other Characteristics of Algae and Algal Groups
Algal groups also differ in the presence of storage products, in cell-wall characteristics and in the fine structure of their cells. Subdivisions of the algae are still the subject of much debate, because of a continuous stream of new information on fine structure and biochemistry.
Algal classification
|
Kingdom: Division: Class: Kingdom: Division: Class: Division: Class: Class: Class: Division: Class: Class: Class: |
Monera (Prokaryota) Cyanophycota Cyanophyceae = cyanobacteria or blue-green algae Protoctista Rhodophycota Rhodophyceae = red algae Chromophycota Bacillariophyceae = diatoms Dinophyceae = dinoflagellates Phaeophyceae = brown algae Chlorophycota = green algae Chlorophyceae Ulvophyceae Charophyceae |
NB:Several other groups of algae exist, but these will not be discussed in the present overview.
Algal Groups
Cyanophyceae are photosynthetic bacteria that are considered algae because their ecology and morphology are rather similar to photosynthetic eukaryotes. Their morphology ranges from unicellular to filamentous, and many possess extensive mucilaginous sheaths. Cyanophyceae occur in marine, freshwater, and soil habitats in a variety of environments. They are planktonic, benthic or free-living and occur in a wide range of temperatures, from ice flow to hot springs. Many form symbiotic associations with plants or fungi, in the latter case often as lichens. Planktonic species can cause water blooms in eutrophic environments, often resulting in health hazards to humans and livestock. Several planktonic and benthic species have the ability to fix atmospheric nitrogen (i.e., nitrogen fixers), which makes them important inhabitants in all marine habitats where nitrogen is scarce, especially in tropical soils (e.g., rice fields).
Rhodophyceae are represented by the macroalgae. However, unicellular representatives and freshwater species also occur. Morphology is rather diverse, ranging from unbranched filamentous forms to crusts, sheets or elaborately branched and relatively highly structured thalli. In the cell walls of some species, commercially important phycocolloids are produced. Calcification of cell walls occurs in several groups, especially in calcareous Corallinales. These calcareous crusts are important consolidators of the frame work of tropical reefs and also produce a part of the reef sediment. Coralline algae, along with the scleractinian corals, are the two dominant groups of reef-builders on Recent coral reefs.
The Chromophycota include all algae possessing chlorophyll-c in addition to chlorophyll-a. Accessory pigments, and especially the brownish xanthophylls, give the chloroplasts their characteristic colours. In fact the Chromophycota embraces a highly heterogeneous group of organisms.
The Bacillariophyceae are the best known and most abundant of all the unicellular planktonic microscopic algae. Diatoms lack the flagella, and possess a box-like silica wall or frustule. The frustules fossilize readily, forming extensive deposits of commercially valuable diatomaceous earth. Diatoms have a wide distribution throughout marine and freshwater environments. They are benthic, planktonic and epiphytic on other organisms.
The Dinophyceae are exclusively unicellular, usually with two flagella. The majority of dinoflagellates are motile, but some are nonmotile. Most members are covered by a wall formed of stiff cellulosic plates. Dinoflagellates are primarily planktonic in both marine and fresh water; some are symbiotic or parasitic. A few of them (e.g., Pyrodinium bahamense var. compressum) form highly toxic water blooms, such as the feared red tides, fish kills and paralytic shellfish poisoning (PSP). A number of benthic reef species (e.g., Gambierdiscus toxicus, Ostreopsis siamensis, Prorocentrum lima) are also responsible for ciguatera poisoning throughout the tropics. Gambierdiscus toxicus is usually associated with benthic brown seaweeds and other macrophytes. The potent neurotoxin (i.e., ciguatoxin) accumulates up the food chain in large predators such as snappers, groupers, and other large reef fish.
Dinoflagellates are symbiotic with a great variety of organisms. Perhaps the best example is Symbiodinium (Gymnodinium) microadriaticum, or the zooxanthellae, which are endosymbiotic with a great variety of reef organisms from sponges to molluscs. There are at least five different taxa of zooxanthellae. These are the algal endosymbionts in corals and giant clams, which facilitate the high photosynthetic productivity responsible for the growth of biotic reefs in tropical waters. However, dinoflagellate symbionts are not restricted only to the benthic environments, since planktonic foraminiferans and radiolarians also possess various photoendosymbionts.
The Phaeophyceae are almost exclusively benthic marine forms. Morphology ranges from microscopic, branched filamentous forms to parenchymatous macroalgae. In this group one finds the highest degree of anatomical differentiation known to occur in algae. Unicellular species are not known. One of the constituents of brown algal cell walls is alginic acid, which is of considerable commercial value. Chlorophycota share with the land plants the occurrence of both chlorophyll-a and chlorophyll-b as main photosynthetic pigments. The vast majority of green algae inhabit freshwater habitats; only 10% are marine. The cell wall consists primarily of cellulose, xylan and mannan. Chloroplasts contain pyrenoids, which are apparently involved in starch synthesis and storage.

Figure 21.4. Gracilaria harvest from coastal brackishwater fish pond.
Photo courtesy of W.F. Prud'homme van Reine
The Class Chlorophyceae contains a variety of unicellular algae as well as colonial, filamentous and parenchymatous ones. The Chlorophyceae are mainly a freshwater group, but a few are essentially terrestrial, living on soil, wood, or leaves. Members of the nonmotile unicellular genus Chlorella are often cultivated as a potential source of health food. Dunaliella species, motile unicellular algae from salt lakes, are used to produce carotenes on a commercial scale.
Ulvophyceae are primarily marine, although freshwater representatives also occur. The thallus of some Ulvophyceae consists of only a few cells, while others are filamentous. Some groups form flat sheets while others are coenocytic. Algae in this group (viz. Caulerpa lentillifera) are used as vegetables, sold fresh or dried. Some genera have calcified walls, viz. Halimeda. Disintegrated calcified Halimeda segments form a significant portion of the white carbonate sand, so characteristic of tropical coral islands.
Charophyceae occur primarily in freshwater habitats. They consist of unicellular, few-celled, filamentous and parenchymatous genera, and are considered more closely related to the land plants than the other classes of green algae.
Seaweeds in Mariculture
Trono and Ganzon-Fortes (1988) published a wealth of information on seaweed resources, seaweed production and the seaweed industry. Of the long list of economically important seaweeds, many also occur in Indonesian waters. About 88 of these usable algae occur in the coastal and offshore waters off Ujung Pandang, South Sulawesi, but only 10 species are used in some form by the coastal people.
The red algae are the most important seaweeds in Indonesian mariculture, and are mainly cultivated for phycocolloids. Along sheltered coastlines species of the genera Eucheuma (mostly E. denticulatum, the "spinosum" of commerce) and Kappaphycus(mostly K. alvarezii and K. striatum, both known as "cottoni") are cultivated on floating rafts or in special seaweed farms, using the monoline method. The phycocolloids produced are different forms of carrageenans. Gracilaria species growing in brackish fish ponds are among the main sources for production of agar, which is another phycocolloid (fig. 21.4). Other seaweeds with economic potential are either produced from wild stocks or are only locally used.

Figure 21.5. Didemnum molle on the move. While considered as a sedentary species, D. molle (Didemnidae) has developed an interesting means of limited locomotion. The animal sends out thin, white fibers, which attach to the substrate some distance from the colony. Once firmly attached, the colony pulls itself along the "anchor lines" to the new position.
Photo by Tomas and Anmarie Tomascik.

Figure 21.6. A sperm release by Aplysina archeri on a Jamaican reef slope at a depth of 53 m (time 15:00 hrs).
Photo courtesy of H.M. Reiswig, McGill University, Montreal, Canada.
Porifera: The Sponges
The Indonesian sponge fauna is among the most species-rich in the world, and is relatively unknown outside a small group of sponge taxonomists (tables 21.2 and 21.3). Most early collections of sponges, made during the various expeditions, were from deep-water habitats. With the advent of scuba, thousands of new specimens have been collected, but, due to the lack of sponge taxonomists, they remain unidentified. Coral reefs are prime sponge habitats. Reef sponges are found in a wide range of habitats, from the exposed outer reef slopes to deep crevices and sheltered caves. Their depth range extends from the intertidal to great depths (fig. 21.6).
Many shallow-water sponges that are present on Indonesian reefs are also common in other areas of the Indo-west Pacific. Van Soest (1985) reported that 26 species appear to be common in northeast Australia, 37 in central Pacific reefs, and 24 in the western Indian Ocean reefs. During the Snellius-II Expeditions, out of the 300-400 species of shallow-water sponges identified from the eastern parts of the archipelago, 56 species were considered common to the area. In addition, a record of the first sclerosponge (Astrosclera wileyana) was made at a depth of about 10 m near Taka Garlarang (Taka Bone Rate Atoll). Van Soest (1985) reported a distinct depth zonation in the shallow-water reef sponge fauna, as well as clear habitat preferences by certain species. For example, 20 species showed a clear preference for sheltered environments, while 23 species preferred exposed habitats.
Table 21.2. Preliminary tabulation of Indonesian demosponge fauna: Class Demospongiae. Totals for taxon in last row.


Table 21.3. Preliminary list of Indonesian genera of glass sponges, Class Hexactinellida (Hyalospongiae), with number of species registered at the Zoological Museum Amsterdam. Totals in last row.

Box 21.2. Indonesian sponges: ecology and biogeography.
R.W.M. van Soest, Institute of Systematics and Population Biology (Zoologisch Museum), University of Amsterdam, Amsterdam, the Netherlands.
Sponges
Sponges, Phylum Porifera, are sessile aquatic organisms with a worldwide distribution that date back to the early Cambrian (560 Ma B.P.) (Fagerstrom 1987). Sponges are a major suspension-feeding invertebrate group inhabiting various aquatic environments from tropical shallow-water marine habitats, such as coral reefs (colour plate 21.2) and anchialine lagoons (colour plate 21.3), down to the deepest ocean trenches.
While most extant sponges are marine, a few hundred species live in freshwater environments. Porifera is a primitive group of invertebrates exhibiting a low grade of cellular organization with a few specialized cells that serve a variety of life-sustaining functions, however, these specialized cells are not organized into tissues or organs. Even though sponges have a simple body structure, they possess a rather complicated internal water canal system that continuously pumps water through the body, thus bringing in oxygen and food as well as removing waste products. Filtration of microscopic-size organic food particles occurs within the water canal system. The only body parts recognizable in all sponges are a "skin" of flattened cells which covers the outside of the sponge, the internal water canal system, and microscopic chambers lined with flagella-bearing cells which generate the water currents for the filtering activity. Sponges also support a great variety of symbiotic organisms, from microscopic prokaryotes to large organisms such as shrimps and fish, which thrive inside their bodies.
Even though sponges lack organ systems, their bodies are supported by mineral or organic skeletons that are secreted by specialized cells. Sponge skeletons consist of discrete siliceous or calcareous elements (spicules) and/or organic collagenous fibers (spongin).
Sponges come in a great variety of shapes and sizes, from flat cushions to elaborate branching, or cup- to barrel-shaped forms (fig. 21.7). The surface of sponges is perforated by numerous microscopic openings called the incurrent pores, as well as a few larger openings called the excurrent oscula. While sponges exhibit great interspecific variation in shape and size, there is also some intraspecific variability which is dependent on environmental factors such as hydrodynamic conditions, light and turbidity.
The knowledge of sponge biodiversity is still largely incomplete. To date, about 70008000 species have been described (van Soest 1994), but as many as twice that number are thought to exist (Hooper and Levi 1994). Sponges are of great interest to science because a large number of species produce chemical substances that have strong anti-viral, antibiotic, anti-fungal and anti-tumor properties. Increasing efforts are being made to screen more species for the presence of these substances. Traditionally, a limited number of species have also been used as "bath" sponges, but these have been largely replaced by synthetic sponges.
Ecology
Sponges are an important component of Indonesian coral reef communities as well as other shallow-water or deep-water marine habitats. In marine ecosystems, major functions are ascribed to sponges. Sponges filter large quantities of organic material from the surrounding water. Each day, a quantity equal to a 40-m-deep and 500-m-wide water column may be filtered by the sponge population of a reef (Reiswig 1974). Sponges compete with other benthic reef organisms (e.g., corals and algae) for space (fig. 21.8) (colour plate 21.4).
In shallow clear-water habitats sponges are less successful, but in more turbid or deeper environments, like mangrove systems or deep reef slopes, they may outcompete reef-building organisms, thus preventing the establishment of a carbonate platform.Specialized sponge genera (e.g., Cliona and Aka) excavate calcareous substrates by chemically etching away tiny pieces of CaCO3. This activity has a great impact on coral reefs and is a major contribution to the recycling of reef carbonates.

Figure 21.7. Sponges can be massive barrel-like organisms (Petrosiasp.) capable of filtering enormous quantities of water, thus, they are often referred to as the reef's filters.
Photo by Tomas and Anmarie Tomascik.

Figure 21.8. Non-zooxanthellate coral Tubastraea sp. being overgrown by an unidentified white sponge in a cryptic habitat.
Photo courtesy of Ron and Valerie Taylor.
Sponges are toxic to many generalized reef predators, and only a few specialized molluscs, echinoderms, fish and one species of marine turtle (Eretmochelys imbricata) are known to feed exclusively on sponges (colour plate 21.5). It is believed that sponge biomass in most habitats is not limited by predation. Wilkinson and Cheshire (1989) demonstrated that biomass of sponges decreases with distance from sources of terrestrial runoff, suggesting that sponges may be limited by food supply. Several specialized genera harbour high quantities of green microsymbionts which act as a source of organic material for the sponge; these sponges are, in fact, "autotrophic", and they occur typically in shallow clear-water habitats.
Indonesian Sponge Biodiversity
The Indo-Malayan region, of which Indonesia is the central part, is the richest marine area of the world (Briggs 1987). Recent reviews of the Indonesian sponge fauna (van Soest 1989, 1990, 1994; Amir 1992; Hooper and Levi 1994), suggest that sponges are no exception to this general pattern. However, the exploration of the sponge biodiversity of the region is still in its initial stages. Only a few older studies on Indonesian sponges (Kieschnick 1896, 1900; Topsent 1897; Thiele 1899, 1903; Vosmaer and Vernhout 1902; Vosmaer 1911; Hentschel 1912; Ijima 1927; Burton 1930; Brodsted 1934), and not a single recent monograph, except for a revision by Desqueyroux (1987), are available to the scientist. Major collections, notably those made by the Siboga Expedition (1899-1900) (approx. 3000 specimens) and the Snellius-ll Expedition (1984-1985) (approx. 1500 specimens), still await description. These collections are housed in the Zoological Museum of Amsterdam. Sponge fauna of adjacent regions (e.g., Papua New Guinea, north Australia and the Philippines) is also poorly known, although huge collections have been made recently and await description (Hooper and Lévi 1994).
The limited information available so far, allows only a few general conclusions about the nature and history of the Indonesian sponge fauna and its affinities to the fauna of other regions. There is an unexpectedly high dissimilarity in sponge faunas of various localities investigated in Indonesia (van Soest 1989; Amir 1992), which means either that we do not know the fauna well enough, or that there is a high degree of endemism within the Indonesian region. With regards to the former reason, there is a personal argument: in my explorations of Indonesian reefs repeated sampling in the same locality invariably yielded species not collected on previous occasions. For the latter reason, for example, the existence of east Indonesian and west Indonesian areas of endemism, there are several geographical and historical arguments (Michaux 1991). An estimated 43% of the sponges recorded from Indonesia appear to be endemic to the region (van Soest 1989). However, this figure is based on "paper" comparisons with faunal lists of other areas, and it is likely that many of the species seemingly shared with other areas will be proven to be slightly different (Hooper and Levi 1994). A number of Indonesian species (5%-10% of the total known so far) occur in many other adjacent areas, often as widely distributed as the Red Sea and the Hawaiian Islands. They are associated with exposed shallow-water coral reefs, and it is assumed that they have mechanisms for maintaining gene flow over large distances (Hooper and Levi 1994). The affinities of the Indonesian sponge fauna are closest with adjacent regions: central Indian Ocean (van Soest 1989; using a biogeographic similarity study) and the central Pacific (Hooper and Levi 1994; using a cladistico-biogeographic analysis). Somewhat surprisingly, affinities with north Australia appear in both studies to be more distant, due, perhaps, to lack of knowledge, but possibly because of Australia's separate Tertiary history (Michaux1991).
One of the most unique discoveries during the Snellius-II Expedition was a symbiotic relationship between the octocoral Tubipora musica and a sponge species of the genus Mycale (Porifera, Demospongiae, Poecilosclerida) (van Soest and Verse-veldt 1987). This previously unknown association was found in the shallow waters around Komodo Island in East Nusa Tenggara. Both organisms are intimately intertwined to form characteristic groups of tubes or pipes (van Soest and Verse-veldt 1987). This new symbiosis is apparently similar to associations between sponges and macroalgae common throughout the Indo-west Pacific and in some places in the Caribbean.
Crustaceans: Pontoniinae
The crustaceans are one of the most abundant groups of invertebrates in Indonesian waters. A large volume of literature exists on this group of organisms whose taxonomy has undergone numerous revisions. Because of limited space we have chosen the Pontoniinae to illustrate the diversity of this fascinating group of marine organisms.
Box 21.3. Indonesian pontoniine shrimps.
Charles H. J.M. Fransen, National Museum of Natural History, Leiden, the Netherlands.
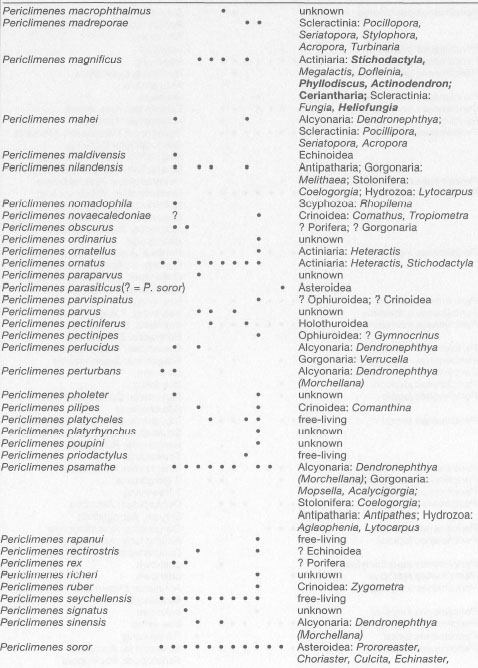
Within the Caridea, the pontoniine shrimps (Crustacea: Decapoda: Palaemonidae: Pontoniinae) are the most radiated subfamily in the marine environment. Of the 420 recognized species, 348 species have been recorded in the Indo-Pacific region (fig. 21.9) (table 21.4). The pontoniids are mainly inhabitants of shallow-water habitats, however, species continue to be found at greater depths down to almost 2000 m. Pontoniid shrimps are found in tropical, subtropical and temperate regions, reaching their highest diversity in the tropical coral reef environments.
The members of this subfamily are small, not exceeding 5 cm in total length. They are often brightly coloured with an amazing variety of colour patterns. One of the main characteristics of the group is that most species live in association with other organisms. Of the more than 80 genera, only five include free-living species, namely: PericlimenesCosta (1844); Palaemonella Dana (1852); Eupontonia Bruce (1971); Exoclimenella Bruce (1994); and Periclimenella Bruce (1994). The pontoniids are known to be associated with Porifera, Hydrozoa, Scyphozoa, Actiniaria, Ceriantharia, Corallimorpharia, Zoantharia, Scleractinia (colour plate 21.6), Stolonifera, Gorgonaria, Alcyonaria, Antipatharia, Pennatularia, Echinoidea (fig. 21.10), Asteroidea, Holothuroidea, Crinoidea, Ophiuroidea (fig. 21.11), Bivalvia, Gastropoda, Nudibranchia, and Ascidiacea. The specificity of the association varies from species to species. Only a few species are known to be associated with a single host species. Most of the pontoniine species live in association with several closely related host species, but not much is known about the nature of these associations. Many shrimp species seem to benefit from the shelter that the host provides, especially those living inside bivalves, sponges and ascidians. Others find shelter among the stinging tentacles of Actiniaria, from which they operate as fish cleaners.
In the past few decades, the number of newly described Indo-Pacific pontoniids increased dramatically, mainly because of the publications by A. J. Bruce (160 species). Exploration of coral reefs using scuba has made many new discoveries possible. From the rate that new species are being described, we can deduce that many more species have yet to be discovered.
The worldwide distribution patterns of many species seem rather incomplete. However, current distribution patterns probably better reflect the distribution of collection sites, rather than the factual distribution of the species. Most remarkable are the usually wide distributions in the Indo-Pacific. For example, some pontoniids are only known from Kenya and Japan, which indicates a lack of data in the intermediate regions. One of the most noticeable 'white spots' (i.e., lack of data) for many species seems to be Indonesia. Only 114 species have been recorded from the Indonesian Archipelago. If one would fill in the discontinuities in the distribution of the Indo-Pacific pontoniids, the number of Indonesian species would increase to at least 170. This number may actually be much higher, since eastern Indonesia forms the centre of marine biodiversity in the Indo-Pacific. The archipelago is a vast region where the faunas of the Indian Ocean, Australia and the west Pacific are superimposed on the Indo-Malayan fauna.
Several recent publications summarize the knowledge on the Indo-Pacific Pontoniine fauna (e.g., Chace and Bruce 1993; Muller 1994; Bruce 1995). Key publications for the pontoniine fauna of Indonesia are: Holthuis 1952; Bruce 1983; and Fransen 1989. Table 21.4 lists the Indo-Pacific Pontoniinae, with their known distribution and host range, based mainly on the above-mentioned publications.

Figure 21.9. Map of areas for table 21.4.

Figure 21.10. Periclimenes cristimanus on a sea-urchin, Diadema sp. SW Sulawesi, 1994.
Photo by C.H. J.M. Fransen.

Figure 21.11. Periclimenes lanipes on an Ophiuroid. Ambon, 1990.
Photo by C.H. J.M. Fransen.
Table 21.4. Checklist of Indo-Pacific pontoniine shrimps with their known distribution and host range. A-K refer to the areas indicated in figure 21.9. • = recorded in the area; ° = now recorded for the first time, based on material in the collection the Nationaal Natuurhistorisch Museum, Leiden (was Rijksmuseum van Natuurlijke Historie); ? = doubtful record. Host genera are enumerated. When genera are not known, only the group-name is given. As far as possible, the recent generic names are used. New host records are in boldface.








Albatross Philippine Expedition 1907-1910, Part 6: Superfamily Palaemonoidea. Smiths.
Contrib. Zool. 543: 1-152.
Fransen, C. H. J. M. (1989). Notes on caridean shrimps collected during the Snellius-ll Expedition. I. Associates of Anthozoa. Neth. J. Sea Res. 23(2): 131-147.
Holthuis, L. B. (1952). The Decapoda of the Siboga Expedition XI. The Palaemonidae collected by the Siboga and Snellius Expedition with remarks on other species. II. Subfamily Pontoniinae. Siboga Exped. Monogr. 39a (10): 1-253.
Muller, H. G. (1993). Catalogue of the Indo-Pacific pontoniine shrimps: 1-159. Wissenschaftlicher Verlag H. G. Muller, Wetzlar.
Giant Clams: Tridacnidae
The giant clams of the Family Tridacnidae were once very common inhabitants of Indonesian coral reef ecosystems throughout the archipelago. While they are still locally abundant, many of the larger species, such as Tridacna gigas, have become locally driven to extinction on many reefs. This is especially true in the more populated parts of the archipelago. Their decline and increased exploitation are occurring despite the fact that they are a fully protected species by law..
While a great deal of research has occurred on the biology and ecology of tridacnid clams in many Pacific nations, research in Indonesia has been progressing relatively slowly. A notable exception are the efforts at the Hasanuddin University in Ujung Pandang, which runs a small field laboratory in the Spermonde Archipelago. A giant clam hatchery has been in operation for some time and, like in many other places in the tropical Pacific, the hatchery methodology has been tested and has been quite successful. However, very few experimental field studies are being conducted to translate the laboratory techniques into a viable giant clam farming project. Considering the continual increase in the population of coastal communities, giant clam farming and reef seeding is the only means by which many wild stocks can be revived.
Box 21.4. Giant clams (Family Tridacnidae).
R.D. Braley, Aquasearch, 6-10 Elena Street, Nelly Bay, Magnetic Island, Qld. 4819, Australia
Giant clams (Family Tridacnidae) are one of the most conspicuous of the numerous invertebrates found on Indo-Pacific coral reefs. The truly gigantic proportions of the largest species, Tridacna gigas, was the genesis of stories in which unsuspecting divers were trapped in giant clam shells. In the Indo-Pacific region, giant clams have long been a traditional source of food and the use of the shell has been important in some localities. In Indonesia, the shells of the largest species of giant clams were the basis of a relatively new floor-tile industry (terrazzo tiles) which flourished from 1961 to the mid-1980s in Java. Panggabean (1987) shows that the Jakarta trade peaked at 7920 tonnes in 1982 and declined to 3677 tonnes and 300 tonnes in 1984 and 1985, respectively. A major site of terrazzo tile production was Jepara, and it was estimated that 45% of the clams had been harvested from the Karimunjawa Islands to supply this trade (Sya'rani 1986). Usher (1984) indicated that local extinction of the larger species has occurred, particularly in western Indonesia.
There have now been nine species of giant clams described. The two most recently described species are not found in Indonesia. These are Tridacna tevoroa Braley (1990) from Tonga and eastern Fiji and Tridacna rosewateri Sirenko and Scarlato (1991), which was only described from empty shells collected at central Indian Ocean reefs. The other seven species are found in Indonesia. The entire family of extant Tridacnidae is listed in table 21.5.
The species which appears to have its geographic distribution centred in Indonesia is Hippopus porcellanus, the China clam. Data on dead shells found in the islands near Ujung Panjang indicate that it grows much larger than previously known from the Philippines (fig. 21.12).
This family of bivalves tends to survive in clumped distributions which are of significance to the reproductive success of the species. When clams are randomly collected for food by fishermen, clumping groups are easily spotted and nearly all are removed. This effectively cuts the reproductive potential significantly because sparse distributions do not contribute to the gene pool. This is due to the distance combined with the short period of time within which released eggs need to be fertilized to attain good fertilization rates (fig. 21.13). If broodstock clams can be collected from sparse distributions and set in a clump, the chance of reproductive success will be increased, but the site must not be vulnerable to poachers. The success of giant clam mariculture on a mass-scale during the 1980s has given hope to the potential of reestablishing giant clam populations in overfished areas. The successes were initiated by MMDC (Micronesian Mariculture Demonstration Centre, Palau) the ACIAR project (Australian Centre for International Agricultural Research) operating at James Cook University in Australia, the University of Philippines and Silliman University in the Philippines, Fiji Fisheries Division, Tonga Fisheries Department, Cook Islands Ministry of Marine Resources, Tuvalu Fisheries Department, Kiribas Fisheries Department and the University of Papua New Guinea, and the ICLARM project (International Centre for Living Aquatic Resources Management) Solomon Islands South Pacific Office). The development of mariculture of giant clams for meat and shell production will help to reduce all but subsistence fishing.
Table 21.5. Family Tridacnidae. The extant species of giant clams, genera Tridacna and Hippopus.


Figure 21.12. Histogram of shell length and shell height of Hippopus porcelanus (live and dead specimens) taken on March 5, 1994, on the islands of Barrang Lompo and Bone Tambung, Spermonde Archipelago, South Sulawesi.
Data by R.B. Braley and A. Rachman.

Figure 21.13. Release of sperm by the giant clam Tridacna gigas on the Great Barrier Reef.
Photo courtesy of R. Steene.
In Indonesia, a law was passed on 12 January, 1987 (Forestry Ministerial Decree No. 12/Ktps/11), which protects all giant clams. However, the policing of this law is difficult and subsistence fishing still contributes considerably to the demise of the giant clams with Indonesia's increasing population. One notable success in Indonesia was a world record cohort of Tridacna derasa produced at the newly constructed giant clam hatchery on Bar-rang Lompo Island, the field station of Hasanuddin University, Ujung Pandang. There, a batch of 1,950,000 three-month-old juveniles were produced. There are some plans to use some of the clams produced from this cohort to introduce to villages which exist within marine reserves such as Taka Bone Rate and Bunaken, both in Sulawesi. This may help introduce the idea of mariculture of clams as an alternative to collection from the wild and also be used to restock areas where the species is very rare. In such a case, it is advisable to place cultured clams in a random or uniform clumping pattern with some wild specimens of the same species to maintain high genetic diversity when spawning occurs.
The Fish Fauna of the Indonesian Seas
The first scientific report on the Indonesian fish fauna was made by Bleeker (1854) on specimens from east Flores (East Nusa Tenggara), which were collected by a Dutch Army officer, J.CJ. Hellmuth, who was stationed at Larantuka. In 1863 Bleeker published a second report that contained 130 species, including 11 which Bleeker (1854) described as new (G. Allen, pers. comm.). Many of these specimens are now deposited at the National Museum of Natural History in Leiden, the Netherlands. According to G. Allen (pers. comm.), very little collecting activity has occurred in Flores, or anywhere else in Indonesia, since that time.
Pieter Bleeker's voluminous and outstanding ichthyological works were, and remain to be, significant contributions to our knowledge of Indonesian fish fauna. During his 36-year career, Bleeker published 500 papers that include descriptions of an incredible number of new taxa: 406 genera and 3324 species (G. Allen, pers. comm.). Approximately 75% of these published articles were devoted to the Indonesian fauna. Revisions of various groups of Indo-Pacific fish by modern researchers frequently attest to Bleeker's uncanny intuition and astute understanding of natural relationships (G. Allen, pers. comm.).
It is common knowledge that the Indo-Australian Archipelago, comprising Indonesia, Malaysia, the Philippines, New Guinea, and northern Australia, is the most diverse region on Earth for fish, and many other coral reef-related organisms. This collective wealth of species was documented most notably by Weber and de Beaufort in their monumental 11-volume series, Fishes of the Indo-Australian Archipelago, published between 1911 and 1962 (G. Allen, pers. comm.). Surprisingly, there is very little information concerning the biodiversity of specific locations within the archipelago (G. Allen, pers. comm.).
One of the most intensive fish studies in Indonesian waters was conducted in Maumere Bay, Flores, by Rudie H. Kuiter and Gerald R. Allen in collaboration with scientists from P30-LIPI. The list offish from this area has grown to about 1133 species belonging to 366 genera in 94 families (G. Allen, pers. comm.). This is the largest number of fish ever recorded from a single, restricted location. Many of these fish were collated pictorially in a handbook by Kuiter (1992). The faunal composition of Maumere Bay is typical of Indo-west Pacific coral reefs. Table 21.6 provides a summary of the number of genera and species per family of all fish found in Maumere Bay, most of which were collected between 1986-1993 by R. Kuiter and G. Allen (Department of Ichthyology, Museum of Victoria and Department of Aquatic Zoology, Western Australian Museum, respectively). Tables 21.7, 21.8 and 21.9 provide a preliminary list of families and species richness of coral reef fish from three different parts of the archipelago collected by G.R. Allen. The data are, however, not suitable for biogeographical comparisons since the areas and sampling efforts are not strictly comparable. Nonetheless, the data serve to illustrate the high diversity of Indonesian coral reef fish fauna, and the need for more research in this fascinating field of study.
The coral reef fish fauna of the Indonesian Archipelago remains poorly known primarily due to the lack of sampling (other than visual counts) in more remote areas. It is not surprising that new species and genera are being described frequently each time ichthyologists visit new areas. Furthermore, our knowledge on the biology and ecology of the hundreds of coral reef fish species remains poor. There is, as yet, little focused marine ichthyological research in Indonesia, and frequently new discoveries on various biological and ecological aspects of coral reef fish are made by accident. Allen and Steene (1995) have recently made an interesting observation regarding the reproductive behaviour of an apogonid, Pterapogon kauderni (fig. 21.14), found at the Banggai Islands off the east coast of Central Sulawesi. P. kaudernivi2& discovered in 1920 and described by Koumans (1933), and is considered to be endemic to this small archipelago (Allen and Steene 1995). As in many cardinalfish (Apogonidae), and other reef fish (fig. 21.15), the male P. kauderni incubates (in the mouth) the eggs until hatching. Allen and Steene (1995), however, found that in this species, the males also orally incubate hatched young. Oral incubation (sheltering) of juveniles among marine fish has not been reported previously (Allen and Steene 1995). It has been suggested that the apparent endemicity of this species to the Banggai Islands is related to lack of pelagic larvae (Allen and Steene 1995). Endemicity among Indonesian coral reef fish is rather low, considering the hundreds of known species. This may not be that unusual considering that the majority of coral reef fish have a pelagic larval stage with high dispersal potential. There are perhaps about 50-100 endemics in Indonesian waters, but we know so little about general distribution patterns that this is purely speculation (G. Allen, pers. comm.).
Table 21.6. Preliminary list of fish families from Maumere Bay, Flores. Number of genera and species per family given.


Table 21.7. Preliminary list of coral reef fish families from Sangalaki Island and Kakaban Atoll, East Kalimantan. Number of genera and species per family given.


Table 21.8. Preliminary list of coral reef fish families from Komodo Island, East Nusa Tenggara. Number of genera and species per family given.


Table 21.9. Preliminary list of coral reef fish families from Manado and surrounding waters, North Sulawesi. Number of genera and species per family given.

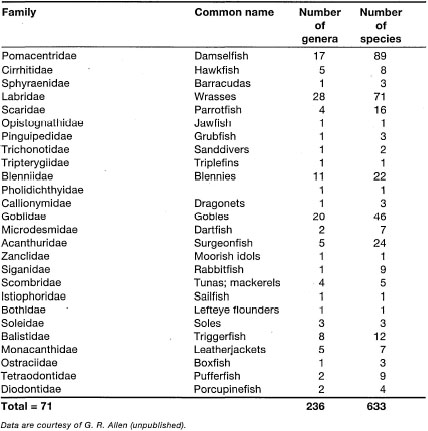

Figure 21.14. Pterapogon kauderni is the only known marine fish to both mouth-brood eggs as well as provide oral shelter for its juveniles after hatching (c. 45 mm SL; Banggai Islands).
Photo courtesy of R. Steene, Cairns.

Figure 21.15. The Indonesian jawfish, Opistognathus sp. (Opistognathidae) mouth-brooding eggs.
Photo courtesy of R. Steene, Cairns.


Figure 21.16. Mutualism on the reef. (A) The "cleaning symbiosis" between Labroides dimidiatus and larger reef fish benefits both the cleaner wrasse which obtains food, and the fish being cleaned, often a reef predator, which gets rid of parasites. (B) Cleaning symbiosis has also evolved in a guild of cleaner shrimps. Lysmata amboinensis attracts fish to its cleaning station by waving its long, white antennae and maxillipeds. The shrimp removes parasites with its maxillipeds.
Photos courtesy of R. Steene, Cairns.
Our knowledge of Indonesian coral reef fish is slowly expanding, however, more ecological and behavioural research needs to be done by the universities. Many coral reefs in Indonesia are close to shore and easily accessible, and universities in Manado, Ujung Pan dang, Ambon, Bali, etc., have relatively easy access to fringing reef habitats. Many ecological and behavioural studies on animal interactions, such as mutualistic relationships (e.g., "cleaning symbiosis") (fig. 21.16) in shallow reefal habitats, require time, dedication and very little else. Many early suggestions about the role of "cleaning stations" still need to be tested. For example, an early hypothesis by Slobodkin and Fishelson (1974) that cleaning stations tend to increase local fish diversity in the Indo-Pacific reefs has not been developed further.
Fish Fauna of the Arafura Sea and Adjacent Waters.
THE MARINE ENVIRONMENT. The Arafura Sea, covering a large part of the vast Sahul Shelf is one of the most important commercial fisheries areas in Indonesian waters. The Arafura Sea covers an area of about 650,000 km2, and includes the Aru Islands and the deep Aru Trough. The north margin of the Arafura Sea is bordered by the Outer Banda Arc to the west and by the southwest coast of Irian Jaya to the northeast. To the southwest, the Torres Strait separates the Arafura Sea from the Coral Sea, while the Timor Sea forms the southwest boundary. The Gulf of Carpentaria forms the southern boundary.
The Arafura Sea covers a vast shallow bank composed of glauconitic sand and calcareous mud (Tjia 1966; Jongsma 1974). The depth over most of the Arafura Shelf varies between 50-80 m. The Aru Trough is 3650 m deep and has an area of 1000 km2 below the 3000 m isobath (Tjia 1966). Surface salinities in die Arafura Sea range between 33.6 - 35 psu (Wyrtki 1961). Maximum average sea surface temperatures occur during December-February (c. 28.4°C), while minimum sea surface temperatures occur during June-August (c. 26.1°C).
The Arafura Sea is under the influence of a strong monsoonal climate. During the Northwest Monsoon (December through March), winds over the Arafura Sea (i.e., between 4°10° S) blow from the west-northwest, with wind speeds of up to 30 km.hr-1. During the Southeast Monsoon (from April through October), southeasterly winds dominate over the area, with wind speeds of about 10-15 km.hr-1.
Sea surface currents north of 8° S have irregular directions and are unsteady, however, south of 8° S the currents during the Northwest Monsoon are predominately westward, running at about 0.25 m.sec-1 (Wyrtki 1961). During the Southeast Monsoon the currents have particular directions in different parts of the sea. In the southwest area of the Arafura Sea, the currents are directed westward into the Indian Ocean through the Timor Sea, while in the southeast area the currents are directed eastward into the Coral Sea (Russell and Houston 1989). The surface current revolves counterclockwise around 10° S and 136° E; speeds do not exceed c. 0.25 m.sec-1. The tidal ranges in the Arafura Sea reach 6 to 9 m at spring tides along the south coast of Irian Jaya and 2.5 m at Dobo, Aru Islands (Russell and Houston 1989). Tidal currents are as much as 0.15 m.sec-1 along the south coast of Irian Jaya, and run in a north-northwest direction, while off the Australian coast tidal currents reach velocities of up to 0.25 m.sec-1 (Russell and Houston 1989).
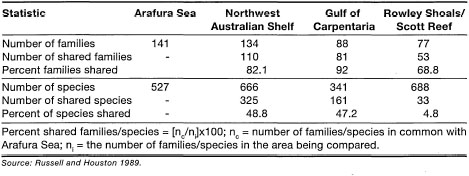
FISHES OF THE ARAFURA SEA AND ADJACENT WATERS. HMS Challenger made the first scientific collection offish from the Arafura Sea in September 1874 (Station 188, 190, 191). They reported 40 species, including 13 that were described as new (Gimther 1880, 1887, 1889). In a study of the Arafura Sea trawl fish fauna, Russell and Houston (1989) recorded a total of 527 species from 141 families. The majority were shallow-water, benthic species belonging to widespread Indo-Pacific families. Other fish were deeper-water or epipelagics with Indo-Pacific or cosmopolitan distributions. The greatest faunal affinity is with the fish fauna of the Northwest Shelf of Australia. The 10 most species-rich families, containing 34% of the total number of species, are: 1) Carangidae; 2) Lutjanidae; 3) Carcharhinidae; 4) Leiognathidae; 5) Nemipteridae; 6) Platycephalidae; 7) Serranidae; 8) Scorpaenida; 9) Mullidae; and 10) Bothidae. In comparison, a total of 146 families and 1140 species offish were recorded from trawl samples in the Indian Ocean waters off the coasts of Sumatra, Java, Bali, and the islands of East Nusa Tenggara during the JETINDOFISH Project (1979-1981) (Gloerfelt-Tarp and Kailola 1984). While the number of families is comparable, the higher number of species collected during the JETINDOFISH Project may be related to high habitat diversity of the area sampled.
A great majority of fish in the Arafura Sea occur in small numbers, and most species individually contribute less than 1% of the mean total catch biomass. Only 23 species are abundant enough in trawl samples to individually contribute more than 1% or more of the mean total catch biomass. These 23 species comprise only 4.4% of the total species and account for c. 70% of the mean total catch biomass of the Arafura Sea demersal trawl fishery (table 21.10).
Comparisons at Family Level. Of the 527 species in 141 families collected in the Arafura Sea, 111 families containing 481 species are shallow-water, benthic forms (i.e., from surface to 100 m). These shallow-water species have a wide distribution throughout the Indo-Pacinc (Springer 1982). There are three endemic families in northern Australian waters (i.e., Tetrabrachiidae, Leptobramidae and Rhinoprenidae) and two families, the Tetrabrachiidae and Leptobramidae, were recorded in the Arafura Sea. In general, the Arafura Sea has a similar number of fish families as the northwest shelf of Australia, but higher than other adjacent waters, even if all deep-water and oceanic families are excluded (table 21.11). The Arafura Sea shares about 50% of the northwest shelf and Gulf of Carpenteria species, but only 30% are shared with eastern Australia (Russell and Houston 1989). This indicates that the Arafura Sea has greater faunal affinity with northern and northwestern Australia than with northern and northeastern Australia. Faunal affinity between the Arafura Sea and the Gulf of Carpentaria is low as a result of the strong influence of the euryhaline species.
Table 21.10. Dominant species taken by trawlers in the Arafura Sea, ranked in order of importance in terms of percent of mean total catch biomass. Only species with greater than 1 % are included.

The comparison of species richness of the 10 most common families in the Arafura Sea indicates that the fish fauna of tropical northern Australia is not homogeneous (table 21.12). The differences are probably related to various environmental and geographic factors which differ among the areas. For example, both the Arafura Sea and the northwest shelf of Australia are continuous open-shelf environments, and, as a result, their dominant fish fauna is characteristic of offshore waters and dominated by carangids, lutjanids, carcharhinids, nemipterids, platycephalids, serranids, scorpaenids and bothids (Russell and Houston 1989).
In comparison, the Gulf of Carpentaria (and Gulf of Papua) contains extensive shallow areas that are subject to wide environmental fluctuations (i.e., temperature and salinity), and the fish fauna comprise a mixture of euryhaline, smidiadromous and marine species (Russell and Houston 1989). Characteristic families of these environmental conditions are carangids, clupeids, scorpaenids, apogonids, leiognathids, ariids, aciaenids, and tetraodontides.
Little similarity exists between the Arafura Sea and Rowley Shoals/Scott Reef, as expected. Some typical reef-associated fish fauna may be associated with sponge/gorgonian-dominated habitats in the Arafura Sea, since similar associations of reef fish with sponge bottoms have been reported elsewhere (Caribbean) (Russell and Houston 1989). Rowley Shoals and Scott Reef are coral reef environments at the seaward margin of the Sahul Shelf, and as a result are very different from the soft-bottom shelf waters of the Arafura Sea, the shallow shelf waters off the north coast of Australia and of the Gulf of Carpenteria. The dominant families are characteristic of coral reef areas, gobiids, labrids, pomacentrids, blenniids, apogonids, serranids, chaetodontids, and acanthurids. An interesting comparison can be made between the Rowley Shoals/Scott Reef and the well-known fish fauna of Maumere Bay in the Flores Sea, since both are coral reef habitats (table 21.13). The Maumere Bay fish fauna is much richer than the Rowley Shoals/Scott Reef fish fauna at both family and species levels. The differences are probably due to higher habitat diversity in Maumere Bay than at the two Australian locations. High habitat diversity in Maumere Bay is reflected by the presence of fringing reefs, barrier reefs, patch reefs, atolls, and various shallow-water coastal habitats, which are, however, not present on Rowley Shoals/Scott Reef due to their offshore location.
Table 21.11. Comparison of Arafura Sea fish fauna with adjacent waters. High number of shared families indicates common faunal origins.
The fish fauna of many tropical continental shelves shows a remarkable consistency, and many of the same families are represented over similar bottom types and similar water masses throughout the tropics (Loghurst and Pauly 1987). The Arafura Sea fish fauna shows strong affinities (in terms offish families) with other areas of the Indo-Pacific, such as the Gulf of Thailand, and the Andaman Sea prior to heavy exploitation (Pauly 1979). The dominant fish families of these areas and the Arafura Sea are typical of fish assemblages which occur in sandy offshore grounds at depths of 50-100 m (Longhurst and Pauly 1987). Species level comparisons are not possible as a result of insufficient data.
The fish fauna of the Arafura Sea has been included in the Dampierian faunal province, an area which extends from the Houtman's Abrolhos (Western Australia) to the Torres Strait (Hedley 1926; Whidey 1932; Bennett and Pope 1953; Knox 1963). However, Endean (1957) questions whether the separation of the Dampierian Province from the Solanderian Province, which includes coastal Queensland, is justified. It is generally recognized that prior to the last Pleistocene flooding, the shelf between Papua New Guinea and northwest Australia must have been relatively isolated7for long periods, leading to faunistic differences (fig. 21.17) (Wilson and Allen 1987). However, various environmental factors, such as the type of bottom deposits, occurrence of coral reefs, water depth, and differences in water masses, are important determinants of species composition of demersal fish assemblages (Russell and Houston 1989). Considering the relatively rapid changes during the Holocene transgression, Russell and Houston (1989) suggested that environmental factors have had, and continue to have, a major influence in determining faunal differences between areas of northern Australia, and that these differences have largely overridden past vicariance events. For this reason they draw short of recognizing distinct zoogeographical regions for northern Australia.
Table 21.12. Composition of the 10 most diverse fish families from Arafura Sea, N.W. Australian Shelf and the south part of the Gulf of Carpentaria.
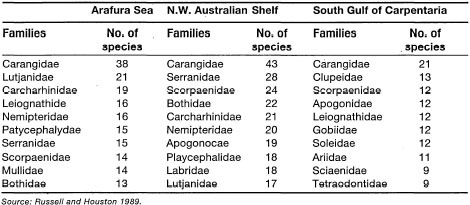

Figure 21.17. The Indonesian Throughflow during the last glacial maximum. Both Sunda and Sahul Shelves were subaerially exposed (heavy line).
From Hantoro 1995.
Table 21.13. Comparison of family species richness in the fish fauna of Rowley Shoals and Scott Reef (Timor Sea) and the fish fauna of Maumere Bay (Flores Sea).

Box 21.5. The damselfish: Indonesia's most abundant family of coral reef fish.
Gerald R. Allen, Western Australian Museum, Perth, Australia.
Introduction
The damselfish of the Family Pomacentridae are one of the largest, hence one of the most important, groups of coral reef fish inhabiting Indonesian seas. Indeed, in terms of number of individuals, no other family of coral reef fish is as abundant. Although they are not large enough to be important for food, some of the larger species of Chromis and Abudefduf are sometimes seen in markets. Because of their sheer weight of numbers, pomacentrids are a vitally important cog in the overall coral reef community. They feed on a wide variety of benthic algae and microinvertebrates as well as a host of planktonic organisms. They, in turn, are preyed upon by larger fish.
Allen (1991) gave a summary of the world's species. He recognized 321 species belonging to 28 genera. Since then, six additional species have been described. A total of 274 species are found in the vast Indo-west Pacific region. Most are tropical dwellers, but the family is also well-represented in cooler seas, for example, along the southern coast of Australia. Indonesia is the world's number one location for damselfish, boasting a total of 138 species.
Damselfish are generally small in stature, seldom exceeding a length of 10-15 cm. Most reef-dwelling species are less than 10 cm and the smallest are fully mature between 4-5 cm. Colouration is highly variable, ranging from drab hues of black, brown, and grey to bright shades of orange, yellow, red, and blue. Many species have juvenile stages characterized by a bright yellow body and with one or more neon blue stripes running back from the head along the upper part of the back. This conspicuous livery gradually fades with growth, often resulting in a nondescript pattern that bears little resemblance to the original colours. The colour pattern for a single species may also vary, although to a lesser degree, according to geographic locality, and more rarely in response to environmental surroundings.
Damselfish are further characterized by a combination of features which include an ovate to elongate and laterally compressed body shape, a single dorsal fin composed of 8 to 17 spines, and a number of flexible, segmented rays, and an anal fin containing two spines and a variable number of segmented rays. The head, body, and fin bases are covered with medium-sized scales that usually have microscopic serrations along the exposed margins. The jaw teeth are arranged in one or two rows and range from a sturdy conical shape to a flattened columnar or spatulate form.
Mainly because of their small size, damselfish have little impact as far as human economy is concerned, except in the aquarium trade. Many damselfish, particularly the colourful members of the genera Amphiprion, Premnas, and Dascyllus, are popular aquarium fish which readily adapt to captivity. Despite their limited economic importance, damselfish play a vital role in the overall food chain of reefs, providing an integral link leading to the larger predators that are often the target of sport anglers and commercial fishermen.
Behaviour
Most damselfish species are highly territorial, or at least are "home-ranging." They swim back and forth over a relatively small portion of reef, often guarding their chosen plot with great zeal, particularly during reproductive periods. Compared with most fish, pomacentrids have highly specialized breeding habits that have been the subject of numerous studies by behavioural scientists.
Several days prior to spawning, the male, or in some cases both partners, selects the nesting site, which is frequently situated under a rock, on a dead coral outcrop, on the inner surface of a dead bivalve shell, or even inside a rusty tin can. The prospective parents prepare the substratum during frequent visits to the nest site. The algal growth is grazed off and debris is removed by vigorous fanning of the pectoral fins. Throughout this period the pair engages in nuptial activities that include chasing bouts, nipping, and rapid side-to-side swimming.
The male may exhibit a peculiar swimming motion consisting of rapid up-and-down movements as though it were on a roller coaster. This behaviour is referred to as signal jumping and serves to attract one or more females to the vicinity of the nest. During the final hours before spawning occurs, these activities are greatly intensified. Eventually the pair skims over the nest surface; the female leads with a slow deliberate motion and exudes a trail of eggs that stick to the bottom. The male follows close behind while fertilizing the spawn. In some species, particularly those of the genera Chromis and Dascyllus, a single male will spawn with two or more females at the same nest.
Spawning often occurs in the morning and may last for more than two hours. Depending on the species, each female lays between about 50 and 1000 or more eggs. Species with larger eggs, such as the anemonefish, have a lengthy incubation period that extends for six or seven days, compared with two to four days in most species. During incubation the male assumes the major role in guarding the nest and in some cases may chase the female from the vicinity.
Nest-guarding males are extremely aggressive and frequently attack much larger fish, especially egg-eating species such as wrasses (Labridae) and butterflyfish (Chaetodontidae). Even skin or scuba divers are not immune from these attacks, which in some species (particularly those in Amphiprion and Stegastes) are accompanied by a stacatto of clearly audible "clicks" or "grunts" and dramatic, instantaneous colour pattern changes. The parents meticulously fan the nest and remove dead or diseased eggs to prevent contamination of the remaining spawn. This process helps keep the nest free of debris and facilitates hatching. The newborn fry vary in length from about 2-4 mm, depending on the species.
Although further research is required, it appears that the young lead a pelagic existence for the first few weeks, drifting near the surface at the mercy of tides, waves, and currents. Judging from the geographic distribution of various species, some may be transported over considerable distances during this stage of the life history.
A recent study (1989) by Thresher, Colin and Bell has resulted in valuable information on the duration of the planktonic larval stage of Indo-west Pacific damselfish. Their study was based on the microstructure of the tiny inner-ear bones (otoliths). They were able to detect daily growth increments during the planktonic or pre-transition stage. Their results indicate that Amphiprion and Premnas have the shortest larval stage, ranging from 7-14 days; Chromis and Dascyllus range from 17-47 days (20-30 days for most); and genera in the Subfamily Pomacentrinae (Abudefduf, Chrysiptera, Pomacentrus,etc.) ranged from 13-42 days. The longest larval periods were detected for Plectroglyphidodon johnstonianus (mean 31.7 days), Stegastes nigricans (mean 32 days), and S. fasciolatus (mean 37.4 days).
The tiny larvae are primarily transparent with scattered pigment cells (melanophores) and silvery eyes. Once swept into shallow inshore reefs, the young fish settle permanently on the bottom and the juvenile colour pattern is quickly assumed. Growth rates of young fish are variable, again depending on the species involved. The rate generally ranges from about 5-15 mm per month, gradually tapering off as maturity is approached. There is very little reliable data concerning the longevity of damselfish, but judging from field studies and aquarium observations, it appears that at least some species may live up to 10 years or more.
Investigations by behavioural scientists working in Japan and the Red Sea have discovered a phenomenal type of sex reversal in anemonefish. Sex reversal of the type in which an individual matures first as a female and later transforms to a male is common among the groupers (Serranidae), wrasses (Labridae), and parrotfish (Scaridae). However, in several Amphiprion species that were studied, the smaller sub-adults possess both male and female gonadal tissue, but only the male testicular tissues are functionally mature. Later in life, at a larger size, these degenerate and the gonads transform into functional ovaries.
Anemonefish are often found in groups of up to five or more individuals and exhibit strict dominance hierarchy or a "pecking order" that is usually related to size. In other words, the smaller fish are chased and generally harassed by the larger ones. Invariably, the largest, dominant member of the group is a mature female and the others are males in various stages of maturity. If the female is eaten by a predator or experimentally removed, the next-largest individual transforms to the female sex within a few days.
Ecology
Although most damselfish occur in the vicinity of coral reefs, they also inhabit a wide range of environmental conditions. At least two Indonesian species, Pomacentrus taeniometopon and Neopomacentrus taeniurus, are frequently encountered in coastal freshwater streams or brackish estuaries. Other species frequent shallow inshore areas including harbours or protected embayments where sandy or silty conditions prevail and visibility is often much reduced. For example, members of the genus Dischistodus are largely confined to this habitat. Most tropical damselfish occur amongst living or dead coral formations in coastal or atoll lagoons or adjacent to drop-offs of the outer reef.
The number of damselfish species present at a given locality is dependent on the variety of habitats that are available. For example, at Christmas Island (Indian Ocean south of Java), where there are no estuaries or protected lagoons, and where conditions are relatively homogenous, the bottom dropping steeply to deep water, 44 species of damsels have been recorded. In contrast, 96 species were seen in the vicinity of Maumere Bay on the Indonesian island of Flores. This huge number is a direct reflection of its rich assortment of habitats.
The number of species found within a specific habitat is directly dependent on the availability of shelter. Therefore, areas where shelter is at a premium - for example, sandy areas, or wave-scoured sections of the intertidal zone or shallow reef crest - have far less species than in those parts of the reef where there is good coral growth (both hard and soft) or other shelter in the form of caves, crevices, boulders or even man-made debris. The importance of shelter is often vividly demonstrated to the diver who encounters swarms of damselfish around isolated coral outcrops or a shipwreck that is situated in a sandy area that is for the most part devoid of fish.
Most tropical damselfish are associated with relatively narrow habitat zones that are best defined in terms of substrata and water depth. Some species, such as Plectroglyphidodon phoenixensis, are restricted to shallow areas exposed to vigorous wave action, while many others, for example, Chrysiptera oxycephalus, frequent sheltered lagoon waters. Amblypomacentrus breviceps and Pristotis jerdoni are usually found in sandy environments, whereas Cheiloprion labiatus is dependent on live coral. Most of the members of the largest pomacentrid genus, Chromis, prefer reefs that are periodically exposed to strong currents that transport their planktonic food supply. The outer reef slope in particular is the domain of many Chromis species.
The great majority of damselfish inhabit depths of less than 15-20 metres. There is, however, a significant number of species that regularly penetrate below this zone. Several Indonesian species, including Acanthochromis polyacanthus, Chromis amboinensis,
C.alpha, C. delta, C. retrofasciata, and P. nigromarginatus, are frequently collected from depths in excess of 60 metres. It is of interest to note that most of these belong to Chromis, a genus that seems especially well adapted for deeper conditions, perhaps because of its planktonic food habits.
The food of damselfish is composed of a wide variety of plants and animals. Some species, for example, those in the genus Stegastes, are mainly herbivorous, feeding on the algal turf that carpets the reef. A number of algal types are consumed, including blue-green algae (Hydrocoleum, Microcoleus, Osillatoria, etc.), green algae (Caulerpa, Chaetomorpha, Cladophora, Enteromorpha, Ulva, etc.), calcareous and filamentous red algae (Jania, Amphiroa, Gracilaria, Ceramium, Polysiphonia, Gel idiurn, Gelidiopsis, Callithamnion, Centrocercas, Galaxaura, Griffihsia, Hypnea, Laurencia, etc.), and brown algae (Dictyota, Ectocarpus, Feldmannia, Padina, Sphacelaria, etc.). The majority of species appear to be omnivorous feeders, although those in Chromis are principally planktivores. The most important animal item appears to be copepods, both benthic and planktonic types. However, a variety of other invertebrate items is also consumed, including siphonophores, corals, sponges, hydroids, tunicates, polychaetes, serpulids, oligochaetes, small gastropods, nudibranchs, ophiuroids, amphipods, isopods, crabs, and shrimps.
Zoogeography
The distribution of a particular damselfish species represents the interaction of several factors. All pomacentrids (except Acanthochromis, see box 21.7) have a pelagic larval stage which varies in length from a few days to several weeks. The infant fish have limited swimming powers at this time and are freely transported in surface waters by wind, waves, and currents. The occurrence of many widespread species (ranging, for example, from the central Pacific to east Africa) that exhibit little or no morphological and colour variation provide good evidence of efficient larval dispersal mechanisms.
Distribution patterns are also dependent on the availability of suitable substrata for the incoming recruits. For example, an anemonefish larva cannot successfully colonize unless the correct species of host anemone is present. Hydrographic features, both past and present, are also influential in shaping distribution patterns. The importance of currents for larval transport has already been mentioned. Heavy discharge of fresh water from rivers and consequent turbidity and siltation may form barriers that prevent larval dispersal, thus limiting the range of a species. Extensive landmasses are also an obvious barrier.
Some researchers have suggested that the Indonesian region, which boasts the richest marine fauna on Earth, represents the evolutionary centre from which the majority of our present tropical reef fish fauna originated. This theory is probably not entirely accurate, particularly in view of the now universally accepted phenomenon of continental drift. In addition, current patterns and climatic regimes were greatly different than those of today. What is certain is that this region has fostered the evolution of the world's most species-rich inshore fish fauna (containing an estimated 3000 species).
This extreme faunal wealth reflects the abundance of marine habitats and microhabitats that are found throughout the Indonesian Archipelago. Some have attempted to explain the huge radiation of species in terms of the ecological stability that has persisted over the ages, thus providing more time for evolution to proceed compared to other regions. However, it seems more likely that the archipelago has experienced a greater variety of catastrophic events (volcanism, sea-level changes and associated emergent land barriers, current shifts, etc.) than other regions, and these vicariant events have created the isolation that is necessary for speciation to occur.
Box 21.6. Inseparable partners.
G.R. Allen, Western Australian Museum, Perth, Australia.
One of the most fascinating aspects of Indonesian marine life is the phenomenon known as symbiosis. This term simply means "living together" and refers to a wide diversity of associations between marine organisms. There are numerous invertebrates that form partnerships that enhance the chance of survival of one or both participants. Various species of crabs and shrimps are among the best-known examples. They frequently live symbiotically with algae, sponges, soft and hard corals, anemones, echinoderms, and ascidians. A number of small fish, particularly gobies, also associate with most of these hosts.
Gobies belonging to several genera (particularly Amblyeleotris, Cryptocentrus, and Ctenigobiops) are invariably associated with alpheid shrimps (fig. 21.18). The shrimp continually excavates the goby's burrow, while the goby keeps a watchful eye for crustacean predators. Keeping an alert vigil at the burrow's entrance, the fish signals the shrimp with a flick of its tail when it is safe to emerge.
Perhaps the best documented example of marine symbiosis involves large tropical sea anemones and damselfish of the genus Amphiprion and Premnas. Indonesian seas are home to all 10 known anemone hosts and 14 fish. Although the host anemones are sometimes seen without fish, there is strong evidence indicating that they are dependent on their fish occupants in order to maintain "peak" condition. The fish remove unwanted debris from the oral disk, and their bathing and grooming behaviour amongst the tentacles is probably beneficial.


Figure 21.18. The shrimp-goby associations. In this mutualistic relationship the shrimp receives protection from the fish which guards the burrow entrance and acts as a lookout for potential predators, while the shrimp builds burrows and tunnels which it shares with the goby. A) Amblyeleotris sp. with Alpheus djiboutensis. (B) Black-rayed shrimp goby, Stonogobius nematodes, with Alpheus randalli.
A. Photo by G.R. Allen; B. Photo courtesy of R. Steene, Cairns.
The fish are absolutely dependent on their anemone hosts for protection. This has been dramatically demonstrated in field conditions by several researchers who have released Amphiprion some distance away from their host and watched them succumb to voracious predators, such as groupers and snappers. The mechanisms that enable the anemonefish to survive amongst the tentacles of its host without being stung have intrigued many researchers. There is now good evidence showing that juvenile anemone-fish acquire their immunity by undergoing a process of acclimation involving very brief contacts with the tentacles. Once fully acclimated, the fish is protected by a chemical additive in their external mucus coat. The fish is not actually immune to the stinging cells (nematocysts) of the anemone, but instead the discharge of the cells is inhibited.
Box 21.7. Coat of many colours.
G.R. Allen, Western Australian Museum, Perth, Australia.
The freshly hatched young of all damselfish, except one, have a pelagic larval stage. The lone exception is the spiny chromis (Acanthochromis poly acanthus). Its spawning habits are typical of the family; a nest of benthic eggs is meticulously cared for by the adult pair. However, the similarity ends at hatching. Instead of drifting away with the currents, the tiny larvae remain in the care of their parents. The adults are very aggressive at this time, summarily driving away any intruders that come near their offspring. Field observations reveal that the fry nip at the mucus coat of the parents, which appears to serve as a source of nourishment, although firm evidence is still lacking.
Individual broods contain an average of about 100 young. As the fry increase in size they become more dependent on the parents for protection and tend to form looser aggregations. After a few months, at a size of about 3-5 cm, the original brood disbands and merges with other groups from adjacent territories.
The lack of a pelagic stage in this damselfish has a profound influence on its overall genetics. In fact, the genetic integrity of the spiny chromis is controversial. Unlike most other damselfish, which have a more-or-less uniform appearance in colour pattern throughout their geographic range, the spiny chromis exhibits a bewildering variety of colour combinations (colour plate 21.7). The explanation for this phenomenon is simple. Because they lack the pelagic stage, individual populations are restricted to islands or reef complexes and are therefore isolated from neighbouring areas. They are incapable of crossing open water barriers.
Thus, a wide array of individual colour patterns has evolved, each characteristic of a particular locality. Each colour variety forms a distinct population that is totally isolated from other populations. Is each population a distinct species? Laboratory examination has failed to reveal any morphological differences besides colour. However, these fish have yet to be investigated using modern techniques that are capable of analyzing genetic composition. Thus far, at least 20 colour varieties have been identified and more will certainly be discovered with further collecting on previously unsampled reefs. The geographic range of the spiny chromis includes Indonesia, the Philippines, New Guinea, New Britain, the Solomon Islands, and northern Australia.
Marine Reptiles
Evolutionary History. There are approximately 6000 species of extant reptiles (Class Reptilia) that evolved from a diverse group of primitive vertebrates during the Mesozoic era (245 - 65 Ma B.P.). Reptiles first appeared in the late Carboniferous (310 Ma B.P.) as an offshoot of a small group of labyrinthodont amphibians (fig. 21.19). This evolutionary breakthrough occurred through the evolution of a new reproductive mode that facilitated the great reptile land radiation during the Mesozoic. The early reptiles acquired the ability for internal fertilization and to lay amniote eggs, in which the developing embryo is enclosed by the amnion, an extraembryonic membrane characteristic of present-day reptiles, birds and mammals. However, in comparison to the great land radiation of the reptiles in the Mesozoic (i.e., Age of Reptiles), only a few groups became specialized for life in the oceans. Among the most specialized were the extinct ichthyosaurs, plesiosaurs and mosasaurs.
In today's oceans, sea turtles (Order Chelonia; Suborder Cryptodira), with sea snakes (Order Squamata; Family Hydrophiidae) and marine lizards (Order Squamata; Family Iguanidae; marine iguana of the Galapagos Islands, Amblyrhynchus cristatus), are the only extant group of true marine reptiles. Sea turtles evolved from the archaic cotylosaurs (stem reptiles) and belong to Anapsida, a subclass of reptiles whose skull lacks temporal fenestrae. During the Mesozoic era sea turtles coexisted with a diverse assemblage of other air-breathing marine reptiles that include the ichthyosaurs and plesiosaurs (fig 21.19). It seems that the environmental changes that may have caused the great extinction of the dinosaurs at the end of the Cretaceous (65 Ma B.P.) did not have a major impact on the evolutionary history of sea turtles.
While the fossil record of turtles is incomplete, it is thought that their evolutionary history dates back to late Triassic time (200 Ma B.P.). The oldest known turtle, Proganochelys quenstedii, dates back to the late Triassic. This extinct group was characterized by the presence of well-developed teeth as well as stout ribs on the neck vertebrae, traits lost in the evolution of modern turtles. It has been recently suggested that pareiasaurs, herbivorous semi-aquatic cotylosaurs from middle to late Permian, are the closest relatives of turtles (Lee 1993). These large armoured herbivores share a number of derived morphological features with turtles, that places them as intermediates (fig. 21.20) between turtles and generalized reptiles (Lee 1993). The recent discovery of a turtle (Kayentachelys aprix) from the early Jurassic (185 Ma B.P.) extends the known record of cryptodires (i.e., turtles able to withdraw their heads into the shells by flexing the neck vertically) back at least 45 million years (Gaffhey et al. 1987). However, the time when the turtles became aquatic is not known. The discovery of Kayentachelys, that has a shell exhibiting morphological features associated with an aquatic habitat, indicates that aquatic habits may have developed early in turtle evolution (Gaffhey et al. 1987). Sea turtles with modern features first appeared in the late Jurassic (140 Ma B.P.) and were common in the Cretaceous (130 Ma B.P.) (Marquez 1990). All extant genera and species of sea turtles originated in the period from early Eocene to the Pleistocene (60 -10 Ma B.P.)

Figure 21.19. Phylogeny of living and fossil amniotes.
A fter Ft a din sky 1987.

Figure 21.20. Phylogeny of living and fossil turtles.
After Radinsky 1987.

Figure 21.21. Phylogeny of living and fossil crocodiles.
A fter Ra dinsky 1987.
Snakes are perhaps the most recently evolved group of reptiles that first appeared during the Cretaceous (about 100 Ma B.P.) as an offshoot of a group of limbless lizards (Heatwole 1987; Radinsky 1987). They underwent a very successful radiation both on land and in the aquatic habitat. Transformations from the ancestral lizard involved the total loss of limbs, increase in body length, increased complexity in intervertebral articulations and of vertebral column musculature, and restructuring of the skull (Radinsky 1987). The Subclass Lepidosauria, to which snakes belong, contains diapsid (i.e., having skull with both dorsal and ventral temporal fenestrae, one above the other) reptiles belonging to one extinct order (i.e., Eosuchia) and two extant orders (i.e., Rhynchocephalia and Squamata). The ancient group of snakes (Family Palaeophidae) survived for about 100 million years and became extinct at the end of the Eocene (c. 35 Ma B.P.). The extant snakes have a wide distribution, ranging from temperate regions to the tropics, where they have achieved their greatest diversification. Their body size varies from 20 cm to over 1000 cm in length. The vast majority of snakes are terrestrial. Snakes (Order Squamata; Suborder Serpentes) are represented by 13 families with about 3000 species worldwide. However, aquatic snakes are represented only in four families (i.e., Acrochordidae, Colubridae, Crotalidae and Hydrophiidae), with Hydrophiidae as the only true marine snakes. The non-venomous Acrochordidae can be found in both estuarine and marine habitats, but are distinct in morphological and physiological features from the venomous Hydrophiidae, and are considered by some as a sister group of the Colubridae (Rieppel 1980).
The present-day crocodilians (Order Crocodilia), consisting of approximately 25 species worldwide, are considered to be the only degenerate survivors of Triassic archosaurians (fig. 21.21). The crocodilians, together with the flying reptiles and the great dinosaurs, are an offshoot of Thecodontia, an order of 'tooth-in-socket' (i.e., thecodont) reptiles that flourished between the late Permian and late Triassic. The crocodilian body plan has changed little during the past 200 million years. All extant crocodilians are aquatic, inhabiting either freshwater or estuarine environments in tropical and subtropical climates. Their main adaptation to the aquatic habitat was the evolution of a laterally compressed tail that serves as the main propulsion organ, and the development of a secondary palate that allows the animal to breathe through its terminal nostrils while the mouth is open under water. Of the four species of crocodiles (Family Crocodylidae) found in Indonesian aquatic habitats (i.e., Tomistoma schlegelii, Crocodilus novaeguineae, C. siamensis, and C.porosus), only the saltwater or estuarine crocodile, C. porosus, ventures into coastal waters. Surprisingly, C. porosus is not particularly specialized for life in salt water, and therefore, it prefers more brackish mangrove habitats.
Sea Turtles.
GENERAL DESCRIPTION. There are approximately 250 species of terrestrial and aquatic turtles worldwide, however, only eight species are marine. Of the eight species of sea turtles found in today's oceans, six species, belonging to two families (i.e., Cheloniidae and Dermochelidae), reside in the Indonesian waters. The most abundant species in the Indonesian waters is the green turtle (Chelonia mydas), followed by the hawksbill (Eretmochelys imbricata), which is found in most coral reef habitats. The demographics of the olive ridley (Lepidochelys olivacea), the loggerhead (Caretta caretta), the leatherback (Dermochelys coriacea) and the flatback (Natator depressus) in Indonesian waters are poorly known. A national sea-turtle survey is urgently needed in Indonesia to obtain accurate demographic information on all species.
With the exception of N. depressus, which is indigenous to the subtropical and tropical Australia, sea turtles have a circumtropical distribution. The six species of sea turtles found in Indonesian waters have a number of common morphological features (figs. 21.22 and fig. 21.23). However, Dermochelys coriacea possesses unique morphological (e.g., leathery, unsealed keeled carapace) and physiological (e.g., chondro-osseus skeleton, well-developed endothermy) traits that place this species in its own family, the Dermochelidae. Some of the key morphological features that the sea turtles share with other cheloniids are: a bony shell consisting of a dorsal carapace and a ventral plastron, which are connected with each other laterally; pendadactyl limbs; jaws modified into a horny beak without teeth; eyelids present; and a non-extensible tongue (Orr 1976). However, the eight species also share a number of common morphological traits that clearly separate them from other cheloniids, namely the presence of nonretractable limbs and head, limbs modified to flippers, flippers with claws, bony skull roof and limited independent movement of the digits.
In addition to morphological adaptations to the marine environment (i.e., presence of flippers and streamlined body), sea turtles also exhibit a number of physiological adaptations, notably the ability to excrete the excess salt ingested with seawater and food, and the ability to dive and stay submerged for long periods. Excess salt (sodium and potassium chlorides) in sea turtles is discharged by means of orbital glands positioned near the eyes. In effect, sea turtles obtain water by drinking seawater and distilling it (Hoar 1966). Sea turtles share this particular physiological adaptation to life in seawater with sea snakes, the marine iguana and sea birds. In addition, ammonia is the major end product in the urine of the green turtle, similar to marine teleosts.

Figure 21.22. Schematic dorsal and ventral views of a sea turtle. C - central (neural) scutes; L lateral (costal) scutes; P - postcentral (pigal, supracaudal, suprapigal) scutes; Pc - precentral scute. 1 - gular scutes; 2- humeral scutes; 3 - pectoral scutes; 4 - abdominal scutes; 5 femoral scutes; 6 - anal scutes.
After Marquez 1990.

Figure 21.23. General structure of sea turtle skull. Pmx premaxilla; Mx - maxilla; Dn dentary; Pf - prefrontal; F frontal; Pa - parietal; Po postorbital; Jg - jugal; Qdr quadrate; Sq - squamosal.
After Marquez 1990.
HISTORICAL PERSPECTIVE. Throughout history, sea turtles have played a major role in the culture and economies of many indigenous coastal populations. Sea turtles were, in the past (Wallace 1869; Wheatley 1959; Sopher 1977), and still are today (Schulz 1984, 1987, 1989, 1992), an important commodity on which many indigenous coastal populations (e.g., Bajau or orang laut) rely for subsistence and trade. The use of sea turtles for subsistence or trade varies from region to region, and is linked loosely to cultural and religious beliefs. Most Moslem communities have a religious aversion to turtle meat (Hendrickson 1958), while a sector of the Balinese population (mainly Hindu) uses turtle meat, not only in certain religious ceremonies (Suwelo and Kuntjoro 1977), but also as a delicacy and a daily staple (fig. 21.24). Irrespective of cultural and/or religious beliefs, turtle eggs are collected throughout the archipelago from most turtle nesting beaches. Increased commercialization in Indonesia has occurred only since the end of the Second World War, with the introduction of motorized fishing boats.
In the past, adat laws and taboos have played an important role in the traditional management of coastal and marine resources by indigenous people. However, because of a rapid population rise and economic development during the past few decades, many of these traditional management techniques were abandoned, or are more-or-less ignored, today. As a result of increasing development pressures, most turtle populations in Indonesian waters are declining (Groombridge and Luxmoore 1989; Schulz 1989), however, no reliable quantitative data, other than declining egg harvests, exist to statistically support this assertion (fig. 21.25). In some areas, harvests of green-turtle eggs have been constant for a number of years, suggesting that the breeding populations have not yet felt the impact (fig. 21.26).
Marine turtles take between 30 to 40 years to sexually mature (i.e., to start laying eggs) (Groombridge and Luxmoore 1989). Sexual maturation rates in turtle populations vary from region to region and are primarily dependent on their diet (quantity and quality), and, to a lesser degree, on climatic conditions (e.g., tropical vs. subtropical; temperature). Late sexual maturation in marine turtles poses serious management implications, since the effects of egg overexploitation will not be felt until the current breeding population of females becomes too old to reproduce. It is theoretically possible to collect 100% of all eggs for the next 20 years and to see no measurable change in the nesting female population numbers and egg production. However, once the population stops reproducing it will crash, since there has been no recruitment of new individuals into the breeding population for the past 20 years. In other words, the current overexploitation of marine turtles may only be manifested 20 years or more from the present, but by that time it will be too late to prevent the collapse of the populations. Because of this unique life history trait, normal fisheries management techniques do not apply to turtles, and must not be used. Ehrenfeld (in Bacon et al. 1984) provides an interesting analogy; he stated that: "Looking at green turtle population data, for example, is like looking at the light from a star 25 light years away: it appears to be shining now, but in fact, you are looking at history, and there is no way of telling whether, during the past 25 years, that star has increased in brightness, or perhaps has gone out altogether".

Figure 21.24. Green turtle (Chelonia mydas) landings in Bali between 1969 and 1989.
Source: Greenpeace 1989; Groombridge and Luxmoore 1989.
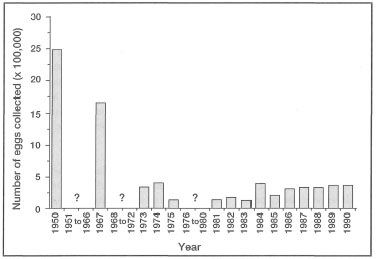
Figure 21.25.Pangumbahan green turtle (Chelonia mydas) egg harvest from 1950 to 1990.
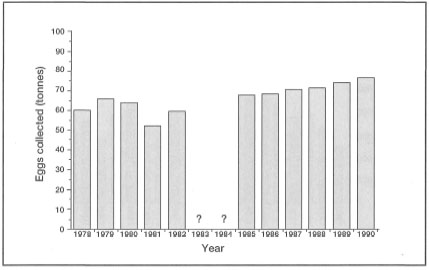
Figure 21.26. Yearly landings of green turtle (Chelonia mydas) eggs in Berau Islands, East Kalimantan, from 1978-1990.
Marine turtles, provided an appropriate conservation plan is implemented, may provide a renewable resource base for subsistence-level use. Because of their unique biology, the large-scale commercial exploitation of marine turtles that we are witnessing today cannot be maintained on a sustainable level. Furthermore, all current research into the biology of marine turtles suggests that attempts to rehabilitate turtle populations through head-starting, farming (breeding), or ranching are not economically viable and are biologically questionable.
The possibility of local extinctions cannot be taken too lightly. Bali is a classic example of local extinction caused by human over exploitation. From a sustainable-use perspective, most egg collectors operating on green turtle nesting beaches throughout the archipelago are destroying a valuable resource, for short-term gain, by removing virtually 100% of laid eggs. Without new recruits into the breeding populations, it is expected that the current green turtle stocks in Indonesia will eventually collapse.
GREEN TURTLE (Chelonia mydas). The green turtle is the largest turtle species in the Family Cheloniidae (fig. 21.27). The streamlined body of the green turtle is depressed in adults, and the carapace is almost oval in shape when viewed dorsally. The width of the carapace is between 85% to 90% of its length. The carapace is smooth and covered by a series of thin flexible scutes. The most noticeable are the five central scutes that are laterally bordered by four pairs of large lateral (costal) scutes. The outer margin of the carapace is covered by 12 pairs of small marginal scutes and a single precentral scute. However, variations in scute numbers have been noted (Forsyth and Balazs 1989). Ventrally, the plastron is covered, along the anterior-posterior axis, by a single intergular scute followed by the paired gular, humeral, pectoral, abdominal, femoral and anal scutes. The terminal scute on the plastron is a single postanal (interanal) scute. The carapace and plastron are connected together by four pairs of inframarginal scutes. The main diagnostic feature that clearly separates this species from the rest of the sea turtles is the presence of two prefrontal scales and four postorbital scales on the head. Each flipper has a single and clearly visible claw. Front claws in males are somewhat larger, and are used to hold on to the females during copulation (fig. 21.28). The length of tail varies depending on sex. In males, tails reach slightly posterior to their hind flippers, whereas in females the tail is very short, barely extending beyond the posterior end of the carapace.

Figure 21.27. Chelonia mydas. Green turtles are one of the most overexploited marine resource in Indonesian waters. C. mydas male (note the long tail) on a patch reef off Sangalaki Island, East Kalimantan, during nesting season (May 1994).
Photo by Tomas and Anmarie Tomascik.
The colouration of green turtles varies worldwide, however, turtle populations in the Indonesian Archipelago show more uniformity. In adult males and females, the carapace is usually deep olive-green with streaks or spots of black, red and yellow. The plastron is usually whitish gray with a yellow tinge to it. C. mydas hatchlings have a black carapace with white margins, while the plastron is white.

Figure 21.28. Chelonia mydas. A copulating pair on a coral reef just off a nesting beach. Note the position of males (top) front flipper. This photo illustrates that green turtles mate during the nesting season in waters adjacent to their nesting beaches. It is essential that they are given full protection in these areas, so that their populations may recover. Sangalaki Island, East Kalimantan, during nesting season (May 1994).
Photo by Tomas and Anmarie Tomascik.
Life History. Green turtles are solitary animals that are, however, known to form large aggregations during breeding periods as well as in feeding areas. Available data on life history characteristics of green turtles have mostly been obtained from their nesting sites (fig. 21.29). While the physiology of the green turtle is well understood, there are still significant gaps in our knowledge with regards to population dynamics as well as on the proximate nature of migrations. All current population size estimates, especially in Indonesia, are approximated from the number of nesting females or egg harvests (Schulz 1984,1987,1989). Information on male abundance and distribution is not well-known.
Both male and female green turtles congregate in waters just off the nesting beaches. Females are frequently found resting on the reef during the day (fig. 21.30), while the males are seen "patrolling" the waters above. The majority of green turtles nest at night, however, in some areas nesting may also occur during early morning hours, just after sunrise (e.g., Sangalaki Island) (fig. 21.29). While green turtles in Indonesia breed throughout the year, there is a well-defined "breeding" season. Most green turtle populations throughout the archipelago exhibit higher nesting activity during certain parts of the year, which differ among different parts of the archipelago (fig. 21.32). In the Indonesian waters, nesting in green turtles seems to be positively correlated with rainfall (fig. 21.33). Lowest nesting activity occurs during the peak dry season, corresponding to higher air temperatures. Comparative studies on nesting seasonality have not been conducted in Indonesian waters.

Figure 21.29. Female green turtle (Chelonia mydas) returning to the sea from nesting early in the morning.
Photo by Tomas and Anmarie Tomascik.

Figure 21.30. Female green turtle (Chelonia mydas)resting on a reef during the day. Sangalaki Island, East Kalimantan.
Photo by Anmarie and Tomas Tomascik.

Figure 21.31. Green turtles (Chelonia mydas) lay eggs mostly at night, however, in some areas, egg-laying during early morning hours is also common. Female green turtle laying eggs on the south coast of Java (time 24:30).
Photo by Tomas and Anmarie Tomascik.

Figure 21.32. Chelonia mydas. A) Seasonal pattern of nesting on Citirem beach, West Java. Nests represent the total number of nests per month. B) Average number of nests laid per night at Sangalaki Island and monthly rainfall.
Data for nesting activity on Sangalaki courtesy of G.F. Taylor, Borneo Divers.

Figure 21.33. Chelonia mydas. Correlation between the monthly average of number of nests laid per night and total monthly rainfall on Sangalaki Island, East Kalimantan.
Data for nesting activity courtesy of G.F. Taylor, Borneo Divers.
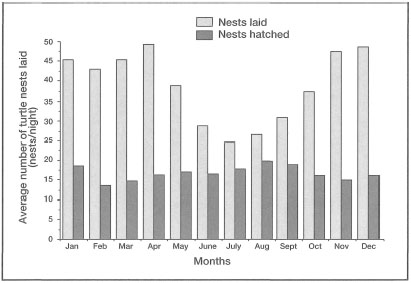
Figure 21.34. Chelonia mydas. Average number of nests laid and the average number of nests hatched per day over a one-year period. Note that while nesting activity has a distinct seasonal trend, number of nests hatched remains relatively steady throughout the year.
Data courtesy of G.F. Taylor, Borneo Divers.
While a considerable amount of data exists in the world literature on both the nesting frequency and clutch size of many green turtle populations, there is very little data available on natural hatching rates. A unique monitoring program was conducted on Sangalaki Island in East Kalimantan, where counts of nests laid and nests hatched were recorded daily. The monitoring program was organized and coordinated by G.F. Taylor (PT Sangalaki Dive Resort). Daily surveys of the entire beach started in April 1993 and continued until July 1994. Based on this extensive database, it appears that only 43% of turtle nests laid eventually hatch (fig. 21.34). The fact that the hatching rate remains relatively steady throughout the year may be related to the carrying capacity of the nesting beach. It is probable that during peak activity (e.g., April and December), a higher number of nests are destroyed by more nesting females, whose population is estimated to be between 4000-5000. This type of information has important implications in the development and implementation of conservation programs. With regards to Sangalaki Island, we may ask the question, 'what happens to the other 60% of the eggs laid?' There is considerable egg predation by monitor lizards and other animals found on the island. However, it is most likely less than 60%. Human predation cannot be excluded, since poaching of eggs was a common affair even when the nesting beach was protected by the resort.

Figure 21.35. Chelonia mydas. A hatchling is being devoured by a large sea anemone Stichodactyla gigantea.
Predation on turtle hatchlings is also intense, however, hatchling mortalities are very difficult to quantify and remain generally unknown. It is believed that high mortalities occur due to predation by large reef predators, such as snappers and groupers. Once the hatchlings are in the open sea, they are presumed to be relatively safe. The journey from the beach to offshore waters is indeed treacherous, for the small hatchlings on Sangalaki Island often fall prey to large sea anemones that occur in great abundance on the surrounding reef flat (fig. 21.35). Whether sea anemone abundance on the shallow reef flat is in any way related to the continuous supply of food (i.e., turtle hatchlings) is not known.
General Distribution. Green turtles have a wide distribution throughout the Indonesian Archipelago, and are considered to be the most abundant sea turtle species found in the region (Groombridge and Luxmoore 1989). Worldwide, there is great variation in the average size of nesting females both in terms of carapace length and weight. This geographical variability in body size is most likely a function of local and regional environmental conditions related to food supply and temperature. The size in all sea turtles is usually measured either in terms of the straight carapace length (SCL) or the curved carapace length (CCL). Because of the slight curvature in the carapace, the CCL measurement is approximately 34 cm longer than SCL (Marquez 1990) (fig. 21.36). Irrespective of the method used, however SCL is preferred, it should always be noted to allow future comparisons. Unconfirmed records from the West Indies suggest that green turtles may reach up to 150 cm in carapace length and weigh up to 395 kg (Marquez 1990).

Figure 21.36. Carapace measurements used in turtle tagging and survey programs. CCL curved carapace length; SCL straight carapace length.
There is little information available on the size distribution of nesting females in Indonesia. However, it is probable that the intense exploitation of nesting females throughout the archipelago, mainly for the Balinese market, has had a measurable effect on the size distribution in recent years. In the heavily exploited Pangumbahan nesting beach, where 95% -1100% of eggs are removed, the average size (CCL) of the nesting females (4 - 9 October 1991) was 102 cm (SD±5.6; N=31), with a minimum of 91 cm and a maximum of 114 cm (Wicaksono 1991). No comparative data are available from areas (e.g., Am Islands) where nesting females are captured for the Balinese market.
Nesting beaches vary in size, location and degree of exposure. For example, the wide beach at Pangumbahan on the south coast of Java is exposed to the swells of the Indian Ocean, while the beaches of Sangalaki Island are sheltered by reefs from most strong wave action (fig. 21.37).
The green turtles have a wide distribution throughout the archipelago, and are generally most abundant in areas with extensive seagrass beds (e.g., Berau Islands, Aru Islands, West Kalimantan, etc.) (fig. 21.38; table 21.14). Their distribution is based on known nesting beaches and feeding grounds. Population numbers of all sea turtles are estimates based on number of breeding females, and are therefore only very rough approximations. Tagging programs on nesting turtles are urgently needed throughout the archipelago to obtain a better approximation. Areas that need to be targeted for immediate research efforts are: Pulau Sangalaki, East Kalimantan; Pangumbahan and Cipatuja, West Java; Pulau Enu, Aru Islands; Pulau Penyu, West Sumatra; Jumbursba-Medi, Irian Jaya; Palok, West Kalimantan.
THE OTHER MARINE TURTLES. The hawksbill turtle is the second-most abundant species of sea turtles in Indonesian waters. It is easily distinguishable from the other species by the presence of thick imbricated (i.e., overlapping) scutes that cover an elliptical carapace. The carapace is covered by five central scutes, four lateral scutes and 11 pairs of marginal scutes. The dorsal scutes are thicker than in C. mydas, and are brightly coloured, which makes them a valuable commodity worldwide. The colouration of scutes varies greatly from region to region. The scutes have radiating streaks consisting of four basic colours (i.e., black, brown, red and yellow) that form fan-like patterns over an amber background. The carapace width is between 70% to 79% of straight carapace length. The hawksbill turtle is taken throughout the archipelago mostly for its valuable carapace, which has long been worked locally. Distribution of hawksbill turtles is shown in figure 21.39. Table 21.15 provides information on population estimates.


Figure 21.37. A) Important green turtle nesting beaches are located along the south coast of Java, where they are exposed to the full force of the Indian Ocean swell. Citerem beach, West Java. B) Important green turtle nesting beaches are also found throughout the sheltered archipelagic seas, where they are protected from the open ocean swell. Sangalaki beach, East Kalimantan.
Photos by Tomas and Anmarie Tomascik.
There is very little information on the ecological function of marine turtles. However, new research in turtle biology suggests that the hawksbill turtle (penyu sisik) may be an important grazer on coral reefs, since it consumes large quantities of marine sponges. It is possible that in certain areas the removal of hawksbill turtles from coral reef ecosystems may eventually result in an overabundance of marine sponges.
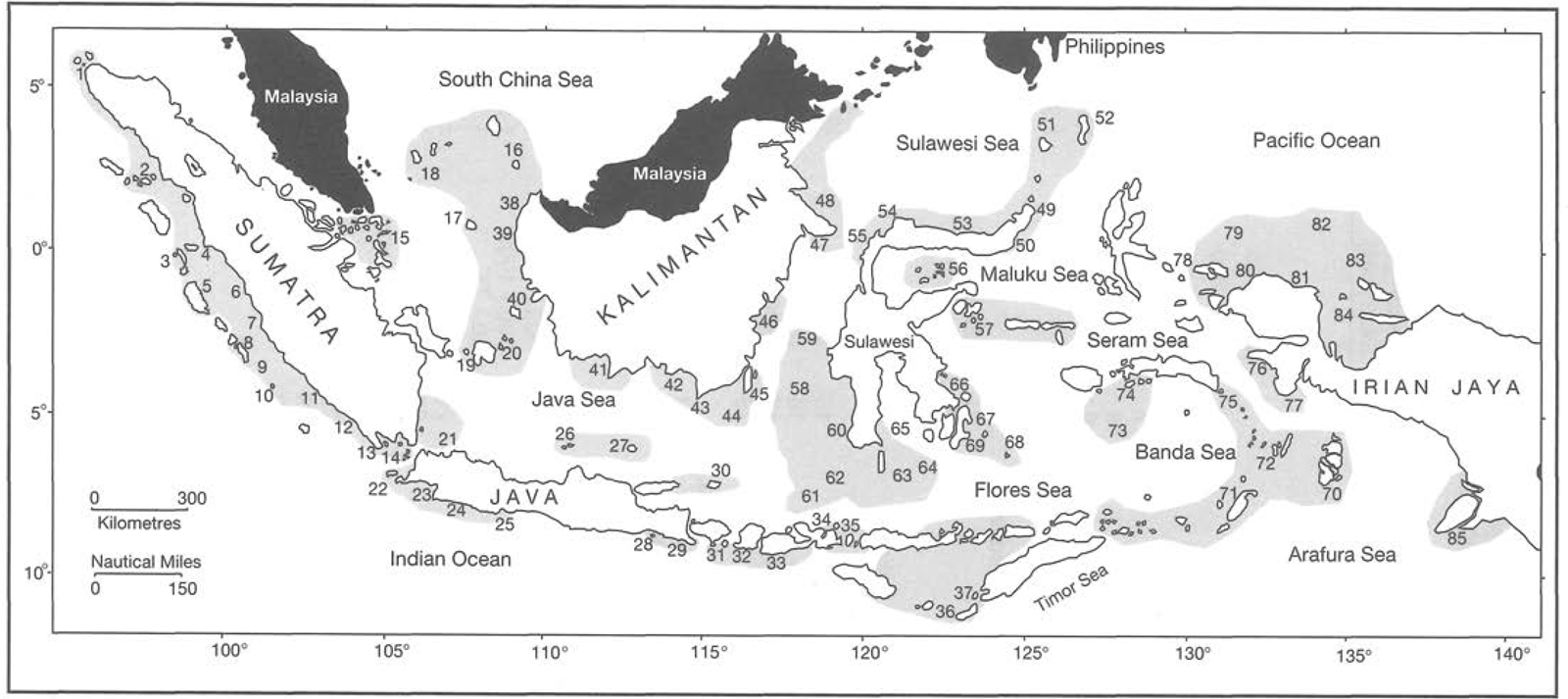
Figure 21.38. General distribution of Chelonia mydas (green turtle) in Indonesian waters. Known nesting beaches are numbered and approximate location is given. Numbers correspond to map numbers given in table 21.14.
Table 21.14. Green turtle, Chelonia mydas, known nesting sites and a yearly estimate of nesting female numbers extrapolated from known egg-collection data (assume 120 eggs per clutch and three or four clutches per nesting season; see Schulz 1987 and Groombridge and Luxmoore 1989). Numbers in the 'Map no.' column correspond to location numbers in figure 21.38.



There are some valuable nesting sites for leatherback turtles (Dermochelys coriaced), although it is rather rare in the intra-archipelagic seas of the region (fig. 21.40; table 21.16). Even though fully protected for some time, the leatherback continues to be hunted in several parts of the country (e.g., Irian Jaya, East Nusa Tenggara); there are important fisheries in the Kai and Aru Islands. One of the major nesting areas of this species is along the north coast of Bird's Head, atjumbursba-Medi and Mubarani-Kaironi beaches.
There is little information on three remaining species, the olive ridley, loggerhead and flatback turtle (figs. 21.41, 21.42, 21.43; table 21.17). There is evidence of a significant decline in green turtle populations connected with sites of heavy meat consumption in Bali, Manado and Ambon. Exploitation of the hawksbill turtle is also heavy due to collecting of untie shells in areas such as Ujung Pandang.
MARINE TURTLE CONSERVATION IN INDONESIA. All six marine turtle species found in the Indonesian Archipelago are currently classified as 'endangered' by the International Union for the Conservation of Nature and Natural Resources (IUCN), and quoted in the Red Data Book. Under the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), to which Indonesia is a signatory, all marine turtles are in appendix I, which stipulates a ban on all international trade in their products. The only trade allowed under the CITES agreement is for turtle products from turtle breeding programs, or for scientific research with a permit from both the scientific and management authorities in the country.

Figure 21.39. General distribution of Eretmochelys imbricata (hawksbill turtle) in Indonesian waters. Known nesting beaches are numbered and approximate locations given. Numbers correspond to map numbers given in table 21.15.
Table 21.15. Hawksbill turtle, Eretmochelys imbricata, known nesting sites and a yearly estimate of nesting female numbers extrapolated from known egg collection data (assume 120 eggs per clutch and three or four clutches per nesting season; see Schulz 1987 and Groombridge and Luxmoore 1989). Numbers in the 'Map no.' column correspond to location numbers in figure 21.39.



Table 21.16. Leatherback turtle, Dermochelys coriacea, known nesting sites and general distribution. Numbers in the 'Map no.'column correspond to location numbers in figure 21.40.


Figure 21.40. General distribution of Dermochelys coriacea (leatherback turtle) in Indonesian waters. Known nesting beaches are numbered and approximate locations given. Numbers correspond to map numbers given in table 21.16.

Figure 21.41. General distribution of Lepidochelys olivacea (olive ridley turtle) in Indonesian waters. Known nesting beaches are numbered and approximate locations given. Numbers correspond to map numbers given in table 21.17.

Figure 21.42. General distribution of Caretta caretta (loggerhead turtle) in Indonesian waters. Known nesting areas are indicated. Figure 21.43. General area where Natator depressus (flatback turtle) is expected to occur.

Figure 21.43. General area where Natator depressus (flatback turtle) is expected to occur.
As a result of new conservation initiatives in Indonesia, two species, namely the hawksbill (E. imbricata) and the flatback (N. depressus), were added in 1992 to the 'fully protected species' list that already includes the leatherback (Dermochelys coriacea), the loggerhead (Caretta caretta) and the olive ridley (Lepidochelys olivacea). The green turtle is the only sea turtle species currently not protected, yet it is widely felt that all green turtle populations in the Indonesian waters are in decline (Soegiarto and Polunin 1982; Schulz 1984). These important conservation regulations are generally ignored by the coastal populations, as well as by many retailers throughout the archipelago where turtle products are available in all major stores. While only Chelonia mydas is systematically hunted, mainly for the Balinese market (for meat), mass collection of all turtle eggs occurs throughout the archipelago, from Aceh to Irian Java. Without strict enforcement of existing laws and regulations combined with a nationwide campaign to conserve turtle populations, this practice will continue and all turtle populations will continue to decline. Local extinctions of green turtles are becoming common, and Bali is a classic example.
The National Strategy and Action Plan for conservation of marine turtles includes the implementation of regulatory measures to reduce the exploitation of green turtles, and gives more protection for species fully protected under existing regulations. Regulations that need immediate implementation are: enforcement of the national green turtle quota (i.e., 3000 green turtles); size restriction, where taking of green turtles with 80 cm curved carapace length or greater is prohibited; full protection of all nesting turtles; and closed season for all egg concession areas.
Table 21.17. Olive ridley turtle, Lepidochelys olivacea, known nesting sites and general distribution. Numbers in the 'Map no.' column corresponds to location numbers in figure

The Action Plan as outlined in the 1992 National Strategy has not been implemented, with the exception of placing the hawksbill and flatback turtles on a protected species list. However, without the implementation of the National Strategy and Action Plan, these new regulations may prove to be in vain.
The Indonesian conservation authorities should review all existing "head-starting" programs (i.e., rearing hatchling turtles in captivity for a certain period of time before releasing them into the sea) in Indonesia, since available information from international scientific research suggests that the practice has not been proven to be an effective management tool. This is essential, since there is a danger that "head-starting" can be used as an excuse for poor management, or, as in the case of Pangumbahan beach, an excuse for no management at all.
Recent evidence points to the possibility that head-starting may not be a sound ecological practice because of: 1) possible interference with "imprinting" mechanisms that eventually guide mature turtles to the nesting beaches; 2) imbalance in sex ratios (recent evidence that artificial incubation of eggs in low temperatures results in all-male broods); 3) nutritional deficiency problems from confined maintenance of hatchlings (we still do not know the food source of juvenile hatchlings in the wild); and 4) behavioral modifications.
Much has been said about the potential of marine ecotourism to support conservation efforts. Yet, if we carefully examine all so-called marine ecotourism destinations in Indonesia, it becomes very clear that the term marine ecotourism has been used indiscriminately, as a successful tourism promotion for many diving destinations. Very seldom do neighbouring coastal communities benefit from marine ecotourism operations, while the reefs are still being degraded. However, there is hope on the horizon, as responsible diving operators are beginning to take conservation initiatives long promoted by the government.
One recent diving destination stands above all others, namely the Sangalaki Island managed by Borneo Divers Ltd. In an innovative conservation effort, the first in Indonesia, the dive operator, PT Sangalaki Dive Resort, bought out the green turtle egg concession on the island, thus effectively putting a stop to egg collection. The people who were employed by the egg concessioner to collect the turtle eggs were now hired by the diving resort to guard the eggs, and thus no jobs were lost. A strict diving code implemented by the diving resort will go a long way to protect the reefs from diver damage. However, the relatively remote location of the destination will ensure that diver or boat damage (anchoring on the reef is not allowed) will be insignificant. Sangalaki Island should be developed as a pilot project where the private sector leads the conservation effort, and where the fruits of their endeavours are spread among the adjacent coastal villages. It should be remembered that ecotourism is much more than just observing nature without destroying it, because it involves people who are, after all, part of nature, a fact that many of us refuse to acknowledge. Ecotourism is a potent conservation tool requiring sound planning and the participation of coastal people.
Conservation Priority Issues.
1)The current exploitation of Indonesian marine turtles and their eggs is proceeding on a non-sustainable basis, which is in direct conflict with the principles of sustainable development.
2) There is a serious lack of fishing regulations governing the commercial and non-commercial use of marine turtles.
3) The use of inappropriate fishing techniques (e.g., blasting and trawling without the turtle excluding devices - TEDs) is having a serious impact on marine turtles and their habitats.
4) Marine turtle nesting beaches are increasingly being degraded by poorly planned coastal development activities and pollution.
5) There is a serious lack of law enforcement of existing laws and regulations governing the commercial and non-commercial use of marine turtles.
6) There is a serious lack of law enforcement of all nesting beaches in protected areas, which results in poaching.
7) There is a serious lack of scientific research on the basic biology and ecology of marine turtles.
8) There is a serious lack of educational material for the general public concerning the conservation of marine turtles.
9) There is a lack of government-sponsored extension programs concerning the conservation of marine turtles.
True Sea Snakes: Hydrophiidae. Sea snakes are the most common marine reptiles in the Southeast Asian seas, and are found in great abundance throughout the Indonesian Archipelago. Along the north coast of Java, south coast of Kalimantan and in the Malacca Strait, snakebites due to sea snakes have been reported among coastal fishermen, but no data are available. In the Arafura Sea, sea snakes are a common shrimp trawler by-catch. The development of antivenins has greatly reduced human mortality rates after envenomation, from roughly 16% to less than 5%. Sea snakes are, however, rather docile animals, and as Gasperetti (1988) points out: "It stretches the imagination to conceive of a sea snake attacking a human being…". On Gunung Api and Pulau Manuk (Banda Sea), thousands of Laticauda semifasciata were found foraging on the reefs, and swimming in the water column over the reefs. They are curious of divers, but can be handled almost with impunity, although this is not a recommended practice, since the venom of this species is among the most toxic.
The true sea snakes belong to the Family Hydrophiidae, which, according to Voris (1977), consists of three major stocks: 1) the Laticauda; 2) the Aipysurus and Emydocephalus; and 3) all other species. Laticauda resembles terrestrial elapids (Elapidae; front-fanged proteroglyphous snakes), and is considered by some workers to constitute the most primitive group of sea snakes (Smith 1926), since it is the least adapted to marine life. Examples of primitive traits are the lateral position of nostrils, development of skin muscles, histology of the skin and oviparous reproduction requiring terrestrial incubation of eggs. Laticauda seems to be intermediary between the terrestrial elapids and the exclusively marine snakes. Aipysurus and Emydocephalus are not as well adapted to marine life as the other Hydrophiidae, while Pelamis is the most adapted to marine life. Enderman (1969) recognized 41 species, which is slightly less than mentioned by others (e.g., 50-55; Gasperetti 1988; Sheppard et al. 1988). However, Enderman's (1969) classification is similar to that of Voris (1977), who recognized 40 species, with several species having one or more subspecies.
The sea snakes are characterized by anterior non-rotatable fangs, neurotoxic venom, lateral compression of the body and flattened oar-like tails (Voris 1977). They have a wide distribution throughout the Indo-Pacific, but are absent from the Atlantic. Even within the Indo-Pacific, their distribution is peculiar. For example, the absence of sea snakes from the Red Sea, the Gulf of Aqaba, and the Gulf of Suez is puzzling, since they are very common in the Arabian Gulf (Gasperetti 1988; Sheppard et al. 1992). It is also notable that only the pelagic sea snake, Pelamis platurus, is found along the east coasts of tropical Africa (Gasperetti 1988). The centre of the geographic range of the Hydrophiidae is the Malacca Strait, where 27 species coexist (Voris 1972). The ecology of the group is poorly known, but they are mainly neritic animals common in mangrove swamps, estuarine bays, river mouths, coral reefs, seagrass beds, coastal mud flats and other coastal habitats (Voris 1972). Only one species, Pelamis platurus, is truly oceanic and is considered as a pelagic species (Tu and Tu 1970). Sea snakes utilize a variety of food types and exhibit high niche diversity (Voris 1972).
ORIGINS. There is no unanimity that sea snakes are monophyletic or a biphyletic group. Most workers agree that their ancestral stock are the terrestrial Elapidae, but probably they have branched off two or more times from the elapids (Mertens 1934; Enderman 1969; Heatwole 1987). The age of the Hydrophiidae remains speculative, since no fossil record of the group has been discovered (Voris 1977). All genera and most species are found in the Indo-Australian seas, between Malaya and New Caledonia. Not much is really known about the evolution of sea snakes, but because they are all poisonous (i.e., proteroglyphous), they may be late arrivals among the serpents. Smith (1926) suggested that genus Laticaudais of Australian origin, while the Hydrophiinae are of Indo-Malayan origin. Others have argued that Laticauda is not closely related to true sea snakes but represents an independent marine adaptation of a group of elapids comprising the Asiatic coral snakes (Calliophis, Maticora), the American coral snakes (Micrurus and Miruroides) and an elapidfrom Bougainville, Solomon Islands (McDowell 1967,1972,1974). However, recent phylogenetic and immunological studies of the group suggest that Laticauda is indeed very closely related to the hydrophiines (Voris 1977; Cadle and Gorman 1981; Mao et al. 1983; McCarthy 1986).
GENERAL MORPHOLOGICAL TRAITS. Sea snakes have evolved special adaptations to the marine lifestyle that clearly make them distinct from their terrestrial relatives, the Elapidae (e.g., cobras). However, even in the Family Hydrophiidae, major differences in morphological, as well as physiological, adaptations occur between the genus Laticauda and the other Hydrophiidae. The characteristic features of Laticauda that make them distinct from the Hydrophiidae are: 1) lateral position of nostrils which requires the snakes to raise their heads slightly above water to breath; 2) a pair of internasal bones on the skull; 3) ventral scales are very broad, 3-4 times the adjacent scale row; 4) subcaudal scales are paired, in two alternating rows; 5) they are oviparous (i.e., lay eggs). In contrast, the characteristic features of the other sea snakes are: 1) superior position of small valvular, crescentic-shaped nostrils with interior valves; 2) internasal bones on the skull are absent, but sometimes pseudo-internasals formed by division of the nasals; 3) ventral scales are broad (up to three times the adjacent scale rows) to very small, divided or absent; 4) subcaudal scales in a single row; and 5) they are viviparous (i.e., give birth to live juveniles). While there are distinct differences among certain groups of sea snakes in general body morphology, they share the following characteristics:
· Body is slightly to strongly laterally compressed;
· Tail short and strongly laterally compressed;
· Poison-fangs followed by 0-18 maxillary teeth;
· Hypapophyses (i.e., a ventral process on vertebra) more-or-less developed throughout the vertebral column;
· Neural spines and hypapophyses very strongly developed in the caudal region;
· Small eyes with round pupils (fig. 21.44);
· Nostrils superior, but lateral in Laticauda;
· Loreal bone absent, but sometimes a pseudo-loreal present;
· Scales strongly imbricate to fully juxtaposed, without apical pits;
· Ventral scales from large to very small to absent;
· Subcaudal scales single or paired.
The sea snakes also share a number of important adaptations to life in the marine environment, which are summarized as follows:
· Adaptation to swimming, strong compression of the body and paddle-shaped tail (fig. 21.45);
· Reduction of ventral scales and development of a keel (i.e., stabilization during swimming);
· Hexagonal scales for less resistance when moving through a fluid medium;
· Reduced length of tongue with only the cleft portion protrusible;
· Superior position of nostrils that can be closed by a spongy tissue - valvular nostrils (not in Laticauda);
· Large right lung extending to the cloaca, where the posterior part serves as a hydrostatic organ and oxygen storage;
· Extrarenal salt excretion is the major route of sodium secretion. Concentrated salt solutions (i.e., brine) are secreted by a gland in the inter-palatine groove - natrial gland;
· Skin permeability; sea snakes have skin quite resistant to the passage of water, but it permits water to move inward more readily than outward. The skin is an effective block to the passage of salt inward (Heatwole 1987);
· Diving adaptation; maximum diving depth of sea snakes is to a depth of about 100 m (Heatwole 1987); the longest submergence time recorded was almost two hours;
· Viviparity except in Laticauda;
· Highly toxic venom delivered by poison fangs present at the front of the upper jaw; the fangs are followed after an interspace by solid teeth posteriorly on the maxillary bone. Teeth are present on the mandible.
Sea snakes also show distinct protective colouration or countershading. Banding in most sea snakes is most intense in juveniles, while less prominent in adults, which may correspond to different lifestyles. In addition, body banding is dimorphic among sexes in most sea snakes. It is interesting to note that a number of eels (e.g., Myrichthys colubrinus) on the reef exhibit Batesian mimicry, and are often confused with sea snakes, most often with Laticauda spp.

Figure 21.44. Laticauda semifasciata. Close-up of the head showing the round pupil. Note the lateral nostrils, which are considered a primitive trait. Manuk volcano, east Banda Sea.
Photo by Tomas and Anmarie Tomascik.

Figure 21.45. Laticauda semifasciata.Compressed, paddlelike tail is a characteristic feature of all sea snakes, which clearly distinguishes them from all other snakes, terrestrial and aquatic. Manuk volcano, east Banda Sea.
Photo by Tomas and Anmarie Tomascik.
Sea snake venom is more toxic than that of their terrestrial relatives. However, because of smaller venom glands, the quantity of venom is much smaller than the amount usually obtained from terrestrial snakes (Tu and Tu 1970), with yields of about 0.5 to 30 mg dry weight per snake (Barme 1963). The venom apparatus in sea snakes consists of the venom gland, the fangs and a connecting duct between the two (Heatwole 1987). The venom in most sea snakes is delivered by two fangs (cobra-like) located on each side of the anterior end of the maxillary bone. Some sea snakes have only one poison fang on each side. The hollow fangs of sea snakes are very short and fixed, unlike in terrestrial vipers which have a complicated mechanism that swings the fangs erect when the snakes bite (Heatwole 1987). Nonetheless, the venom glands and related musculature of sea snakes conform in their general structure to those of the terrestrial elapids (Gopalakrishnakone and Kochva 1990). In species such as Aipysurus eydouxii and Emydocephalus, the fangs are greatly reduced in size, since these sea snakes are primarily egg-eaters. However, they still have small but functioning venom glands (Gopalakrishnakone and Kochva 1990).
Sea snake venom is a complex solution of many different substances (e.g., c. 25 different enzymes) (Heatwole 1987). However, compared to terrestrial vipers, sea snake venom contains fewer enzymes (Tu and Tu 1970). Not all sea snake toxins are enzymes. For example, Uwatoko et al. (1966a,b) reported that the venom of Laticauda semifasciata (common in the Moluccas) contains neither protease nor cholinesterase activity. Tu et al. (1966) found an absence of endopeptidase (i.e., proteinase that splits a protein molecule into smaller units by hydrolysis), thus venoms of some sea snakes cannot hydrolyze (i.e., addition of the hydrogen and hydroxyl ions of water to a molecule which splits it into two or more simpler molecules) certain protein molecules. Different species of sea snakes have different toxins, each with specific effects.
Sea snake venom can be classified as neurotoxins, myotoxins and haematoxins. Neurotoxins (polypeptides) act on the neuromuscular junctions between nerve and muscle cells. Some venoms affect the cell membranes of nerve cells and interfere with the release of acetylcholine, which is required for normal neural transmission. Other neurotoxins allow the release of acetylcholine from the nerve cells, but block its transmission by binding to the muscle cell receptors (Heatwole 1987). The venom of Pelamis platurus is known to have strong neurotoxic action, and 0.5 mg of the venom can kill a guinea pig within two hours by producing asphyxia and paralysis of the respiratory system (Nauck 1929; Tu and Tu 1970). Myotoxins are enzymes that directly break down muscle fibers. The most common are phospholipases, which cause muscle cells to break down into fatty acids and lysophospholipids (Heatwole 1987). One side-effect of direct myotoxin action is the release of myoglobin, which may block renal tubules and eventually cause renal failure (Heatwole 1987). Haematoxins are enzymes that affect blood. Some haematoxins break down cell walls of red blood cells, thus releasing haemoglobin (eventual respiratory failure), while others interfere in clotting properties of the blood (Heatwole 1987). For example, the venom of Laticauda semifasciata produced hemolysis of rabbit erythrocytes (Tu and Tu 1970).
Venoms of Laticauda laticaudata, Enhydrina schistosa, Hydrophis cyanocinctus and Lapemis hardwickii have a strong neuromuscular blocking action in experimental animals (e.g., guinea pigs, frogs, rabbits, etc.) (Carey and Wright 1961; Chan and Geh 1967; Cheymol et al. 1967; Tu 1963, 1967). More recent toxicology studies have used coral reef prey fish as experimental subjects. For example, coral reef fish (e.g., Chromis nitida, Dascyllus aruanus and Istiblennius meleagris) injected with the venom of Aipysurus laevis exhibit six progressive stages of envenomation. The effects of envenomation, defined behaviorally, were common to all three species (Zimmerman et al. 1990; Zimmerman and Heatwole 1992). Depending on the concentration used, the venom caused a rapid decline in control of mouth and ventilatory movements, loss of fin control, swimming coordination and buoyancy and eventual death. The specific effects of the venom on prey fit with the feeding mode of A. laevis, which is a "crevice-forager" (Zimmerman et al. 1990). However, Zimmerman et al. (1992) demonstrated that there are significant differences in response time to the venom among different reef fish species. Apparently, blennies (e.g., Istiblennius meleagris and J. edentulus) have higher survival times under high venom doses than pomacentrids (Zimmerman et al. 1992). In contrast to experimental studies on laboratory animals, sea snake venom is known to produce necrotic lesions in muscle tissue as well as myoglobinuria in human sea snake bite victims (Reid 1956; Marsden and Reid 1961).
REPRODUCTION. Most sea snakes are viviparous - they give birth to live young. The exception is Laticauda, which is oviparous (i.e., lays eggs) and must lay its eggs either on beaches or in caves. Very little information is available on the reproductive seasonality of sea snakes in Indonesian waters. Bergman (1943) studied the reproduction of six species from the Java Sea (i.e., Thalassophis anomalus, Lapemis hardwickii, Enhydrina schistosa, Hydrophis fasciatus, Hydrophis brookii and Hydrophis cyanocinctus). His findings on sex ratios are presented in table 21.18.
In the Java Sea, egg development in female sea snakes begins in March and lasts until August. By September the embryos are well developed, and the actual pregnancy lasts between May to November (Bergman 1943). By the beginning of November most female sea snakes have given birth, indicating that the gestation period in these species is about seven to eight months. The number of young born varies from species to species. Bergman (1943) found that Lapemis hardwickii and Hydrophis fasciatus generally give birth to two young. Thalassophis anomalous and Enhydrina schistosa produce five young, while Hydrophis brookii bears seven. The most fecund species is Hydrophis cyanocinctus, which gives birth to 10 young (Bergman 1943).
Reproductive cycles in sea snakes vary greatly among species and from place to place. In some species reproductive activity is continuous throughout the year, while in others there are clear seasonal cycles (Heatwole 1987). Very little information exists on the reproductive activity of Laticauda in Indonesian waters. The two largest Laticauda semifasciata populations in Indonesian waters were found at Gunung Manuk and Gunung Api in the Banda Sea. These two active volcanic islands support thousands of sea snakes. Numerous caves offer ideal nesting places that are well protected from most predators. Why there are so many sea snakes at these two islands remains a mystery, but it is an exciting area for future research, not to mention adventure.
Table 21.18. Sex ratios in Java sea snakes. Sea snakes from the north coast of Java.

FEEDING HABITS. Sea snakes take a great variety of food types, but many are highly specific for one particular food type. Two or three species of sea snakes are generalists, but most species specialize on eels, goby-like fish or catfish (Voris et al. 1978; Voris and Voris 1983). Lapemis hardwickii and Aipysurus laevis are good examples of generalists, preying on a total of 31 and 12 fish families, respectively (Voris and Voris 1983). In contrast, Ephalophis greyi preys predominantly on Gobiidae, while Emydocephalus annulatus is an egg-eater (Voris and Voris 1983). Like their terrestrial relatives, sea snakes swallow their prey whole. As a result, fish size and shape may be important prey characteristics. Voris (1972) has classified sea snake prey into four body-type categories, namely: 1) eels (e.g., Apodes) and eel-like fish (e.g., Synodus); 2) bullet-shaped fish (e.g., mullets); 3) vertically or laterally oversized fish (e.g., puffers and flatfish); and 4) other than fish (e.g., bird eggs, fish eggs, and prawns). Voris (1972) also classified sea snake prey according to ecological characteristics, namely: 1) inshore, on or near bottom; 2) rock-dwellers; 3) reef-dwellers; 4) pelagic inshore; and 5) pelagic offshore. Voris and Voris (1988) found that the families of eels and gobies are taken by the greatest number of sea snake species. It seems that most sea snakes prefer fish that are bottom-dwelling, relatively sedentary, live in burrows or crevices, and have fine scales.
Foraging behaviour in sea snakes can be classified as: 1) "crevice-foraging"; 2) "cruising near the bottom"; and 3) ambush (Heatwole 1987). Sea snakes, such as Laticauda colubrina, are good examples of species that employ the crevice-foraging method. At Pulau Manuk and Gunung Api, these species were observed to forage during day and night. In this type of foraging activity, the sea snakes move over the reef frequently, protruding their tongues and poking their heads into small fissures and crevices in the reef. The female L. colubrina feeds specifically on the conger eels (Conger cinerus), while the male feeds primarily on echinde eels (Pernetta 1977; Ackman et al. 1991). The reefs of both Pulau Manuk and Gunung Api supported exceptionally high reef fish biomass, including eels. Note that both islands are located in an upwelling area, which may explain the high productivity and abundance of sea snakes.
The second method of "cruising near bottom" is employed by species such as Enhydrina schistosa. This species forages by swimming just above the bottom and when it encounters an acceptable food item it strikes. Most often the strike is lateral, and misses are frequent. Once the prey has been captured, it is held in the mouth until the poison has taken effect. Once the prey has stopped moving, it is ingested headfirst (Voris et al. 1978).
The ambush method is employed by the pelagic species Pelamis platurus. Many small pelagic fish are known to be attracted to floating objects where they seek shelter from predators. The yellow-bellied sea snake (P. platurus) forms small aggregations that resemble floating slicks. Small fish are attracted by these "floating objects" and seek shelter beneath them. Once they approach close enough, the sea snakes strike and ingest the prey headfirst. This pelagic snake is able to swim forward and backward equally well (Heatwole 1987).
GENERAL DISTRIBUTION OF SEA SNAKES. The classification of venomous sea snakes remains problematic at best. The following discussion does not deal with systematics, and the reader should refer to references indicated for the taxonomic treatment of the group. The main objective here is to provide new information on general distribution of sea snakes in Indonesian waters and adjacent seas. Enderman (1969) considered sea snakes to be a biphyletic group consisting of two subfamilies (i.e., Laticaudinae and Hydrophiinae) belonging to the Family Elapidae. Burger and Natsuno (1974) consider Laticauda as unrelated to Hydrophiidae and placed them in the Family Laticaudidae. McCarthy (1985) treated sea snakes as a monophyletic group, and treated them as subfamilies of Family Elapidae. However, the most widely held view is that all extant sea snakes belong to the Family Hydrophiidae (Voris 1977; McCarthy 1986).
Table 21.19 lists all the known sea snake species (modified from Enderman 1969), and indicates which species are found in the Indonesian waters. Note that a number of species have subspecies present. Even with our limited database on the sea snake fauna of the Indonesian Archipelago, the data presented in table 21.19 demonstrate that c. 70% of sea snake species found throughout the Indo-Pacific are found in Indonesian waters. The species so far not recorded in the Archipelago are considered to be endemic to the North Australian Shelf (e.g., Ashmore reefs). However, intensive sampling in the Arafura Sea has not been conducted, and thus their endemicity remains rather questionable.
The following account of sea snakes in the Indonesian Archipelago and adjacent waters is based on the work by Enderman (1969), which one of us (T. Tomascik) had the privilege to review, thanks to Dr. M.S. Hoogmoed from the National Museum of Natural History, Leiden. All new information, unless cited otherwise, is fully credited to Enderman (1969) and the National Museum of Natural History, Leiden. Please note that this is not a taxonomic treatment of the Hydrophiidae.
Family Hydrophiidae.
Subfamily Hydrophiinae.
Genus Aipysurus Lacepede, 1804. Of the seven strictly marine species of Aipysurus, four are found in the Indonesian waters, and the rest appear to be endemic to the north Australian continental shelf area (south Timor Sea). They are known to congregate in large numbers on some reefs. Females produce less than four young per breeding season, however, the young are quite large (Cogger 1975).
Aipysurus eydouxii (Gray). This species is found from Southeast Asia to New Caledonia and Queensland. It is very common in Thailand, South Vietnam, Singapore and north Java, but it has not been recorded in the Philippines. The species is apparently absent from the Malacca Strait, which is known to support 27 species of sea snakes (Voris 1977).
In Indonesia, and adjacent waters, this species has been recorded in Singapore, Jakarta, Cirebon, Semarang, Pasuruan, Probolingo, and other places in the Java Sea. The species is very common along the north coast of Java, where it is found in shallow coastal waters, sandy shores and river mouths. Fishermen in Cirebon and Probolingo are very afraid of this species, which is interesting, since it is primarily a fish egg-eater with greatly reduced fangs and a low volume of venom. A. eydouxii has also been reported from Sarawak, and therefore, it may occur along the coastline of Kalimantan. Three species are endemic to Australian waters, but only A. laevis and A. duboisii are widely distributed in that region. Their limited distribution range suggests specific ecological requirements. They are apparently not able to live and/or cross the muddy seas of the Sahul Shelf, while the deep seas in the Moluccas may be another barrier. A. eydouxii does not normally inhabit coral reef areas and does not occur with Laticauda, however, it may be found in areas where these habitats exist together, (e.g., Singapore, N.W. Borneo). The distribution of several sea snake species along the east coast of Southeast Asia - from the Strait of Malacca to Tonkin Bay - is interesting but cannot be explained. The interrupted distribution may be caused by ecological conditions.
Their general habits are poorly known. Apparently they are gentle and do not bite unprovoked. They are known to move fast but awkwardly on land, as was observed on specimens from Surabaya (Flower 1896). Aipysurus spp. do not leave water.
Aipysurus fitscus (Tschudi). The type specimen of this species is from "Celebes" (Sulawesi). However, in a three-year study in the Arafura Sea and adjacent waters, Shuntov (1972) failed to collect any specimens other than those from the reefs off the north Australian continental shelf margin (e.g., Ashmore reefs).
Aipysurus tenuis (Lonnberg and Andersson). Aipysurus tenuis has a limited distribution in the coastal waters off northwest Australia (Cape Jaubert near Broome) and in the Arafura Sea (Shuntov 1972).
Table 21.19. The list of sea snakes of the Family Hydrophiidae (44 species). Sea snakes collected in Indonesian waters are marked with '+'. Species not recorded in Indonesia are marked with '-'.

Aipysurus laevis (Lacepede). Aipysurus laevis is distributed along the north coast of Australia, from Broome (Western Australia) to Rockhampton (Queens land) to New Caledonia. In Indonesia it has been recorded from the Kai Islands, the Aru Islands and the Arafura Sea. The species is rather common in Australian waters on the Sahul Shelf. It is also generally common on the Great Barrier Reef with Laticauda colubrina.
Genus Astoria Fischer, 1855
Astoria stokesii (Gray, 1846). Astoria stokesii is a large, heavily built sea snake that reaches up to 1.5 m in length. The broods are small, with less than five young produced per breeding season. It is distributed from west of the Indian Peninsula (Malabar coast and Sri Lanka) throughout the Southeast Asian seas (Singapore; Gulf of Thailand) and to all waters of tropical Australia.
A. stokesii seems to occur in three disjunct areas: 1) west of India; 2) Natuna Sea. (e.g., Singapore); and 3) Australia. It has not been found in any intermediate waters. Since it seems to be absent from Madras as well as the Java Sea, the three populations are probably reproductively isolated. The Malayan form from the Natuna Sea has higher body band numbers, and occurs in the Strait of Malacca, Singapore, the Gulf of Thailand and Patani Bay. Reports suggest that this species forms large migrating groups (in the millions) in the Strait of Malacca. These may also represent breeding congregations. The Australian form extends from northwestern Australia to the Aru Islands to New South Wales, Australia.
Genus Kerilia Gray, 1849
Kerilia jerdonii (Gray, 1849). Not much is known about this monotypic genus. Its distribution is from south India to South Vietnam. In Indonesian waters it has been recorded along the west coast of Borneo, in the Malacca Strait (Penang, Malaysia) and in the mouth of the Pontianak River. Two subspecies are recognized, namely K. j. siamensis and K. j. jerdonii.
Genus Kolpophis Smith, 1926
Kolpophis annandalei (Laidlaw, 1901). This species ranges from the south coast of Vietnam through the southern parts of the Gulf of Thailand to the north entrance of the Malacca Strait (Patani Bay). It is not very common in the Java Sea, however, it apparently is locally common in the mouth of the Mengala River (Lampung, Sumatra).
Genus Lapemis Gray, 1834
Lapemis curtus (Shaw, 1802). This species has a wide Indo-west Pacific distribution from the Arabian Gulf to North China and south to tropical Australia. In Indonesia, it is found in Aceh, Siak (Sumatra), the Natuna Sea, the Java Sea (e.g., Jakarta Bay, Cirebon, Pekalongan, Semarang), the Strait of Madura, Sumba, Flores, Sulawesi (e.g., Ujung Pandang, Manado) and the Moluccas.
Two subspecies are present, but they are considered allopatric: L. c. curtus and L. c. hardwickii. Both subspecies are very successful snakes and often locally the commonest snakes in the coastal waters. L. c. hardwickii is widely distributed, and its presence in the Mergui Archipelago (off the east coast of Myanmar), but its absence from the east coast of India, suggests that there is no reproductive isolation with regards to curtus (there is no more formidable barrier for a subspecies than another subspecies). L. c. hardwickii has two forms; an Indo-Malayan form and a Philippine form. The Philippine form was described from specimens collected in Borneo and Sulawesi.
Genus Praescutata Wall, 1921
Praescutata viperina (Schmidt, 1852). Praescutata viperina is widely distributed from the Arabian Gulf to Shantou (China), but it is absent in the Philippines. In Indonesia it is present in the Malacca Strait, Java Sea (e.g., Semarang, Surabaya), West Kalimantan (e.g., Pontianak) as well as in the Sunda Strait.
Genus Thalassophis Schmidt, 1852
Thalassophis anomalus (Schmidt, 1852). This species has a wide distribution in Southeast Asian waters. Its northern distribution extends from the northern part of the Gulf of Thailand to Sarawak (Malaysia). Its southern distribution extends from East Sumatra (Siak) to the Java Sea (north coast of Java), the Strait of Madura and to the Moluccas. T. anomalus has a highly disjunct distribution. While common in the north Gulf of Thailand, it is virtually absent from the adjacent waters, and while T.anomalus is common in east Sumatra, it has not been recorded in intermediate waters.
Genus Pelamis Daudin, 1803
Pelamis platurus (Linnaeus, 1766). This pelagic species has the widest geographic distribution of all true sea snakes, extending from the east coast of Africa to the coast of Baja, California. Its latitudinal range extends from Possiet Bay in Siberia to Tasmania. It is the only truly pelagic sea snake species. It is found in most Indone sian intra-archipelagic seas.
Genus Enhydrina Gray
The genus extends from the Arabian Gulf to tropical Australia as far as Rockhampton (Queensland), and northward to South Vietnam, Sarawak, Mindanao, and the southwest coast of Irian Jaya.
Enhydrina schistosa (Daudin). This is a somewhat problematic genus. Many workers consider Enhydrina to be a monotypic genus, but McDowell (1972) synonymized it with Disteria, which was resurrected from Hydrophis. Here Enhydrina schistosa is regarded as a single species with four subspecies. Disteria is synonymized with Hydrophis.
Enhydrina prefers shallow, sandy seas and estuaries, and is absent from the rocky shores of Sri Lanka, the Philippines and the Moluccas. It is the most frequent sea snake found in rivers in Sumatra. Fairly common in Digul River (Irian Jaya), up to 450 km from the river mouth. Stomach contents consist of pufferfish and moray eels.
The four subspecies recognized are: 1) E. s. schistosa which extends from the Arabian Gulf to the Mergui Archipelago; 2) E. s. smithii extending from the Malacca Strait to the South China Sea and into south Indochina; 3) E. s. werneri found in the Java Sea (Java); and 4) E. s. digulis found in the waters of New Guinea.
The Irian Jaya population differs from the Asiatic populations. Java specimens differ from Malayan specimens by having two postoculars (62%) rather than the usual one (79%-90%). Enderman (1969) treats the Java specimens as distinct subspecies.
Specimens from northeast Sumatra resemble the Indian form, however, specimens from the Sunda Strait and south coast of Sumatra more closely resemble the Java form. The Gulf of Thailand population resembles the Java Sea population.
It was suggested that the Java Sea (Java) population may have arisen from a hybridization between the populations occupying the South China Sea and along the east coast of the Sunda Shelf during the last glacial. The recent populations of the Gulf of Thailand and the Madura Strait represent the direct descendants of both glacial populations (Enderman 1969).
The E. s. werneri (Schmidt, 1852) subspecies is considered to be the Java Sea population (form), with specimens from Semarang. The population is distributed mainly along the north coast of Central Java. Even within the north Java population morphological differences are apparent (table 21.20).
The Enhydrina schistosa digulis subspecies is found mainly along the southwest coast of Irian Java, from Digul River southwards to Merauke. The freshwater and sea populations may be different subspecies, but sample size is not large enough to make a meaningful comparison.
Genus Kolpophis Smith, 1926
Kolpophis annandalei. This poorly known species occurs in the Malacca Strait, extending to South Vietnam, but is absent from the north part of the Gulf of Thailand. It is confined to shallow muddy coastal waters.
Genus Hydrelaps Boulenger, 1896
The genus contains two species distributed along the north coast of Australia and south coast of Irian Jaya and Papua New Guinea. However, Cogger (1975) considered it a monotypic genus. It has been fully redescribed by McDowell (1972), who considered it to be the most primitive sea snake.
Hydrelaps darwiniensis (Boulenger, 1896). This species is found from Port Darwin to Merauke in Irian Jaya, but little is known about it.
Hydrelaps mertoni (Roux, 1910). This species is represented by a single specimen from Sungai Waskai, Wokam Island, Aru Islands. Burger and Natsuno (1974) have erected a new genus, Parahydrophis, to accommodate this species. However, here we shall follow the classification of Enderman (1969). Cogger (1975) noted the work of Burger and Natsuno (1974), but chose to follow the classification proposed by McDowell (1969, 1974), who described the species as Ephalophis mertoni.
Genus Hydrophis Latreille, 1803
This is the largest genus of sea snakes, with about 20-25 species, depending on classification used. The exact number is unknown. McDowell (1972) recognized 21 species. Here we discuss 17 species and five subspecies as suggested by Enderman (1960). The genus has a wide distribution and its members are found in a variety of habitats. Cogger (1975) points out that members of this genus are subject to marked ontogenic variation in colour, pattern, morphology and allometry, which greatly complicate its taxonomy, especially when sample size is small. Very little is actually known about each species in Indonesian waters.
Table 21.20. Geographic variation of some morphological features (in %) within the subspecies Enhydrina schistosa wernei. A) Prefrontals touching labials; B) Fourth only at eye; C) Cuneate present; D) Two postoculars present; E) Number of specimens examined.

Hydrophis belcheri (Gray). McCarthy and Warrell (1991) have recently described Hydrophis belcheri from the Java Sea (e.g., Karimunjawa Islands and south coast of South Kalimantan) and the Gulf of Thailand. Its distribution extends to the coastal waters of Irian Jaya, but not much is known about it and the species is known from only a few individuals. Tamiya and Puffer (1974) regarded the venom of H. belcheri as one of the most potent they have analyzed. McCarthy and Warrell (1975) noted that one of their female specimens contained four young.
Hydrophis spiralis (Shaw, 1802). Hydrophis spiralis extends from the Arabian Gulf to the Ryukyu Islands (Japan), Fiji, and New South Wales (Australia). H. spiralis prefers rocky shores to estuarine habitats, contrary to most members of this genus. While it is very common in the Ryukyu Islands and tropical Australia, and fairly common in the Moluccas, it is rare in the Java Sea and absent in the Gulf of Thailand, and along the Indo-Chinese coast and South China Sea. There are four subspecies recognized, but only H. s. spiralis and H. s. brugmansii occur in Indonesian waters. H. s. spiralis is restricted to the Malacca Strait and the Riau coastline, while H. s. brugmansii has a wide distribution from Malaysia to Brunei and the Philippines, south to New Guinea and southwest to the Pacific. It is common in the Aru Islands, Babo (Bintuni Bay), Waigeo Island, as well as in Tanimbar, Timor, Sumba, Morotai and Wotap Islands.
Because of its abundance throughout the Moluccas and in the Pacific (Gilbert Island, New Caledonia, Fiji), it may be able to bridge deep seas, and therefore, may be more pelagic than most sea snakes, with the exception of Pelamis platurus.
Hydrophis elegans (Gray). Very little is known about H. elegans in Indonesian waters, even though it is an abundant species in Australian waters and along the south coast of Papua New Guinea. Cogger (1975) indicated its presence in the coastal waters of Onin Peninsula and Salawati Island in eastern Irian Jaya. Females may contain up to 23 embryos (Cogger 1975).
Hydrophis melanosoma (Gunther, 1864). Hydrophis melanosoma is a little-known sea snake, found in the Malacca Strait from the coast of Perak to East Sumatra. Its distribution range extends northeast across the Natuna Sea to northwest Borneo (e.g., Brunei Bay). H. melanosoma has a rather limited distribution within this area, and is absent in the Gulf of Thailand and Indochina. Enderman (1969) suggested that H. melanosoma evolved from H. spiralis during the Pleistocene period by geographic isolation, since H. spiralis does not occur in the South China Sea, while H. melanosoma is only known from the Malacca Strait and northwest Borneo. The current H. melanosoma population may exist in reproductive isolation.
Hydrophis cyanocinctus (Daudin, 1803). H. cyanocinctus occurs in the Moluccas but is relatively rare. It extends into the southwest Pacific and tropical Australia. It is quite common from the Arabian Gulf to the west coast of India (Bombay), through to the Malacca Strait, the Gulf of Thailand and northward to Hong Kong. H. cyanocinctus is also common in the Java Sea and extends northward to the Philippines (e.g., Manila Bay).
Hydrophis klossi (Boulenger, 1912). Hydrophis klossi is found throughout the Malacca Strait, along the coasts of Perak and Selangor. The species extends north ward to the Gulf of Thailand. H. klossi consists of two isolated populations west and east of the Malay Peninsula.
Hydrophis torquatus (Gunther, 1864). The geographic range of Hydrophis torquatus extends from the southeast coast of India (Madras) westward to Borneo and northward to south China. However, H. torquatus is absent in the Philippines and in the Java Sea. Its Malacca Strait population extends through the Natuna Sea into the South China Sea. This species has been recorded in the Gulf of Thailand (Taylor 1965). Enderman (1960) recognized three subspecies, but only one which occurs in Indonesian waters. H. t. torquatus is found throughout the Malacca Strait and along the east coast of Sumatra northward to Banda Aceh. This group inhabits shallow coastal waters and estuaries, and is known to venture up rivers.
H t. torquatus has a limited range, which suggests that it has very specific eco logical requirements, which may explain its absence from Java. Its apparent preference for estuarine habitats may account for its absence from Singapore.
Hydrophis caerulescens (Shaw, 1802). Hydrophis caerulescens is found from the south coast of Pakistan (Karachi) eastward to Java, but it is absent in the southern part of India and in the Philippines. One specimen is from the Moluccas. H. caerulescens seems to prefer estuarine habitats and shallow coastal seas. It is abun dant in the Gulf of Thailand and along the northwest coast of Borneo (Sarawak).
Enderman (1969) recognized two subspecies. H. c. caerulescens which is found in the Malacca Strait, along the east coast of Sumatra (e.g., Riau). Its western distribution extends from the Mergui Archipelago to Karachi (Pakistan). H. c. hybridusis found in the Java Sea (Banten Bay, Cirebon, Semarang, Surabaya, Probolingo, Pasuruan), the Sunda Strait, the Moluccas, Borneo (Satubong), the Gulf of Thailand, southeast Vietnam and along the south coast of China.
Hydrophis fasciatus (Schneider, 1799). Hydrophis fasciatus is widespread from Pakistan (Karachi) eastward to south China (Gulf of Tonkin), east to Philippines and south to northern Australia. Two subspecies are recognized, with the Riau Archipelago being a boundary between them. H. f. fasciatus is the western group found from Karachi to Singapore. It is extremely rare, west of the Indian subcontinent, and no records exist for Sri Lanka. H. f. fasciatus is very common between Madras and Sunderbands (Bangladesh). It has been recorded along the west coast of Myanmar (Akyab and Rangoon). It is relatively common in the Malacca Strait. H. f. atriceps is the eastern group, extending throughout Southeast Asian waters to northern Australia. It has been recorded in the Gulf of Thailand, South Vietnam, the Gulf of Tonkin, and in the Philippines. In Indonesia, it is found in Kalimantan, Java, Sulawesi (Ujung Pandang) and the Aru Islands. H. f. atriceps can be locally very abundant.
Hydrophis ornatus (Gray, 1842). Hydrophis ornatus is widely distributed from the Arabian Gulf to north China (Tsingtau), the Philippines and south to Tasmania. In Indonesian waters, H. ornatus is quite common in the Java Sea along the north coast of Java. According to Enderman (1969), there are three possible subspecies, with one present in Indonesian waters. H. o. ornatus is found from the Arabian Gulf to the Ryukyu Islands, Japan. In Indonesian waters some records exist from Java and the Moluccas. However, it is generally rare or absent from the shallow seas on the Sunda Shelf, since it prefers rocky coastlines without muddy rivers. Enderman (1969) points out that it is able to cover relatively long distances over open ocean, and that it resembles H. spiralis and its subspecies.
Hydrophis major (Shaw, 1802). Hydrophis major has a limited distribution throughout tropical Australia, from Broome (W. Australia) to New Caledonia. In Indonesia it is common along the southwest coast of Irian Jaya, from Merauke to the mouth of the Digul River. Cogger (1975) treats this species as Disteria major (Shaw), following McDowell's (1972) classification in which he resurrected Disteria from the synonymy of Hydrophis. There is a strong colour-pattern similarity between H. major and Astoria stokesii. H. major appears to be restricted to deeper turbid coastal waters, and is taken by shrimp trawlers in the Arafura Sea.
Hydrophis lapemoides (Gray, 1849). Earlier distribution of Hydrophis lapemoides extended from the Arabian Gulf to the east coast of India, and in the coastal waters of Sri Lanka (McCarthy and Warrell 1991). However, the species has recently been recorded from the southern part of the Malacca Strait and the Gulf of Thailand (Rasmussen 1987; McCarthy and Warrell 1991).
Genus Microcephalophis Lesson, 1834
The genus has two species, one found in Indonesian waters. McDowell (1972) relegated genus Microcephalophis to the synonymy of Hydrophis, and placed gracilis in subgenus Hydrophis. For the sake of continuity we choose to follow Enderman's (1969) original classification.
Microcephalophis gracilis (Shaw, 1802). Microcephalophis gracilis is widely distributed from the Arabian Gulf to south China. It is relatively common in the coastal waters of Borneo, and along the north coast of Java. Its southern distribution extends to tropical Australia. However, M. gracilis is absent in the Philippines.
Three subspecies are present, each with marked geographic variation, but only one has been recorded in Indonesian waters. M. g. microcephalics extends from the Mergui Archipelago through the Malacca Strait (e.g., Perak and Selangor) and to northwest Sumatra (Singkilbaru, Aceh). From the Malacca Strait, M. g. microcephalics extends westward through the Natuna Sea and along the north coast of Java to the Madura Strait (e.g., Probolingo). The Java and Malayan forms differ in the number of ventral scales and may be two subspecies, however, the sample size of specimens from Java is not sufficient to make a definite conclusion.
Subfamily Laticaudinae. The relationship of Laticaudinae to other sea snakes (Hydrophiinae) remains confused. Some herpetologists consider genus Laticaudaas being the most primitive group of sea snakes belonging to the Family Hydrophiidae (Smith 1926). In his phylogenic analysis of the Hydrophiidae, Voris (1977) concluded that the genus Laticauda is a phenetically tight group of closely related species, and that it has a weak phyletic relationship with a number of Aipysurus and Emydocephalus (Hydrophiinae) species. Voris (1977) suggested that: "…these groups may have independent origins among the elapids or a single origin with a very early separation." Others have, however, suggested that Laticauda (Laticaudinae) and the true seas snakes (Hydrophiinae) have had separate origins within the Family Elapidae (McDowell 1967, 1969, 1972, 1974; McCarthy 1985).
Genus Laticauda Laurenti, 1768. Laticauda is widely distributed from the Bay of Bengal (Calcutta) to the Society Islands (Tahiti) in the central Pacific. Its latitudinal distribution extends from southern Japan (Tsu-shima in the Korea Strait) southward to Tasmania and New Zealand. The genus is closely associated with coral reef ecosystems as well as rocky shores of tropical and subtropical environments, and is the sea snake most often seen by divers. Laticauda is considered as an incidental visitor to Sunderbands and the north parts of New Zealand. Three species are found throughout the Indonesian Archipelago (table 21.21).
Simple Key to Species of Genus Laticauda,
|
1. Upper lip yellow 1. Upper lip brown 2. No azygous prefrontal; rostral undivided 2. An azygous prefrontal; rostral divided |
Laticauda colubrina 2 Laticauda laticaudata Laticauda semifasciata |
Laticauda laticaudata (Linnaeus, 1758). Laticauda laticaudata occurs throughout the archipelago, but seems to be more common in the eastern regions. Colouration of L. laticaudata exhibits ontogenic changes. The bluish black body bands are very intense in small specimens (< 50 cm). Larger individuals (50-100 cm) are bluish grey to olive green or greyish above, yellowish below, tail with yellowish tip. Males have between 4-7 tail bands, while females have 2-5 tail bands. The body may have 28 to 66 black or dark-brown bands. In larger specimens (>100 cm), the banding fades and the rings are darkish brown in colour.
L. laticaudata is widely distributed from the Bay of Bengal to Ryukyu, Tonga and Tasmania. It inhabits tropical and subtropical coral reef environments and rocky coastlines. While having a similar distribution as L. colubrina, it does not occur in either Singapore or the Riau Archipelago.
Laticauda colubrina (Schneider, 1799). Laticauda colubrina has a rather similar distribution as L. laticaudata and on many reefs they occur together. This species is found from Aceh to Irian Jaya and from North Sulawesi to Sumba. L. colubrina can be easily distinguished from the other two Laticauda species by its bright yellow upper lip. The body bands in L. colubrina are darker in colour than in L. laticaudata and do not seem to fade with age as much. The tail bands in L. colubrina show regional variability (table 21.22).
Laticauda semifasciata (Reinwardt in Schlegel, 1837). The type species of Laticauda semifasciata is from Solor Island, East Nusa Tenggara. It is distinguished from the L. laticaudata and L. colubrina by having the rostral plate divided horizontally into a large lower portion which is broader than high, and a small upper portion lying between the nasals. The nostrils are positioned near the middle of the nasal. The dorsum of the head has three prefrontals with very short parietals which are not longer than broad and much shorter than the frontal (fig. 21.47).
The colouration of L. semifasciata is greyish, bluish, or greenish above, yellowish below, with 21 to 43 blackish brown bands on the body, and 4-8 on the tail. With age, bands fade ventrally and become lighter, more yellowish brown. Large specimens are brownish with the markings hardly apparent. The head is dark brown with a curved white/yellow mark above, and connected to the first white/yellow band on the neck.

Figure 21.46. Dorsum of sea Rostral snake head showing the position of major plates.
Modified from McCarthy 1986.
Laticauda semifasciata is found in southern Japan (Ryukyu Islands), the Philippines, the Moluccas, the Lesser Sunda Islands, Irian Jaya (Biak) and the western Pacific. The two largest populations of L. semifasciata in Indonesian waters occur at Pulau Manuk and Gunung Api, Banda Sea. At both locations we observed thousands of L. semifasciata. This species seems to prefers caves for nesting (fig. 21.48). Large aggregations (hundreds of individuals) were seen inside caves, where the snakes were observed to enter into crevices and fissures in the ceiling of the caves. It is suspected that the entrances lead to small caves above the main subtidal cave where the snakes may lay their eggs. Tu et al. (1990) reported that in Taiwan, Laticauda semifasciata lay their eggs in caves where moisture levels during incubation were maintained by fresh water dripping from condensation on the cave roof. During our survey dives, numerous "courtship" behaviours were observed. Furthermore, the snakes in the cave were entangled with both sexes present. It is suspected that these large aggregations may occur during mass-breeding events. Both islands are surrounded by fringing reefs (fig. 21.49). The two islands were visited at the end of the Southeast Monsoon (October) just at the end of the upwelling season, when the waters are rich in zooplankton and the reefs surrounding the two volcanoes support high fish biomass. Both Gunung Api and Pulau Manuk offer exciting research opportunities to study two large sea snake populations.
Table 21.21. Genus Laticauda. Morphological differences between species. Numbers in brackets indicate 'rarely'. See figure 21.46 for location of plates.


Figure 21.47. Close-up of the Laticauda semifasciata showing the arrangement of bone plates on the dorsum of the head. Pulau Manuk, Banda Sea.
Photo by Tomas and Anmarie Tomascik.
Table 21.22. Geographic variation in the number of tail bands in Laticauda colubrina.


Figure 21.48. Laticauda semifasciata at an entrance to a cave on Gunung Api.
Photo by Tomas and Anmarie Tomascik.

Figure 21.49. The fringing reef of Gunung Api, Banda Sea.
Photo by Tomas and Anmarie Tomascik.
Key to the Genera of Hydrophiidae.
1. Nostrils lateral; nasals separated by a pair of internasals and the rostral; ventrals very broad, 3-4 times the adjacent scale row; subcaudals paired, in two alternating rows..... Laticauda
1. Nostrils superior; nasals broadly in contact or separated by pseudo-internasals (in the latter case ventrals are very small); ventrals absent, divided, narrow, or broad, up to three times the adjacent scale row; subcaudals in a single row..... 2
2. Ventrals broad, c. three times as broad as adjacent scale row; scales imbricate, 1723 at mid-body; anterior temporals divided..... 3
2. Ventrals narrow, less than three times as broad as the adjacent scale row, sometimes divided or absent; scales imbricate or juxtaposed..... 4
3. Supralabials 6-10..... Aipysurus
3. Supralabials three, the second very long and touching the eye..... Emydocephalus
4. Mental long and narrow, partly hidden in a groove in the chin; rostral higher than broad, with a downward projection beyond the level of the lip..... Enhydrina
4. Mental small and triangular; chin groove absent; rostral as broad as or broader than high..... 5
5. Anterior ventrals broad, 3-4 times as broad as the adjacent scales, narrowing gradually posteriorly where they are not twice the adjacent scales..... Praescutata
5. Ventrals not or hardly widened anteriorly..... 6
6. Lowermost 3-4 scale rows enlarged; ventrals posteriorly narrow, fragmentary or absent..... Lapemis
6. Lowermost scale rows not enlarged; ventrals present throughout, undivided or completely divided..... 7
7. Rostral divided into 4-5 pieces; nasals separated by a pair of elongated shields..... Thalassophis
7. Rostral undivided, nasals in contact with each other or irregularly divided..... 8
8. Supraoculars and usually some adjacent shields raised posteriorly and ending in a spine; frontal and parietals broken up..... Acalyptophis
8. Head shields not raised and ending in a spine..... 9
9. Scales 62-82 and 74-96, very small and juxtaposed; head shields (especially nasals and prefrontals) irregularly divided....... Kolpophis
9. Less scale rows; upper head shields undivided..... 10
10. Ventrals completely divided into two overlapping scale-like halves; scales strongly imbricate Astrotia 10. Ventrals undivided or divided into two juxtaposed halves..... 11
11. Head elongated; body short, not slender anteriorly, very strongly compressed; scales small and juxtaposed throughout; 39-55 scales on the neck; ventrals usually divided by median furrow, the halves hardly distinguishable from the dorsal scales; distinctive colouration; normally a dorsal black band and a yellow or brown ventral side..... Pelamis
11. Head short; ventrals undivided or divided (divided only in Microcephalopsis, which is very slender anteriorly and has 17-25 scales on the neck); usually a banded colour pattern, sometimes grayish above, whitish below..... 12
12. Snout much narrowed anteriorly; scales 16-19. 19-23 (w), imbricate; Ventrals 212-293 and not much broader than the adjacent scale row..... Kerilia
12. Snout not particularly narrowed anteriorly; on the body scales 19 (only in Ephalophis, which has 165-168 broad ventrals and is endemic to northwest Australia) or 25-59..... 13
13. Body long and slender anteriorly; ventrals entire on the slender part, completely divided posteriorly by a median furrow, the two halves apposed or alternating..... Microcephalophis
13. Ventrals entire throughout (like in most sea snakes incidentally divided), nearly twice or more than twice as broad as the adjacent scale row..... 14
14. Ventrals 160-172, more than twice as broad as the adjacent scale row 15 15. Ventrals 195 - 514 . . Hydrophis 15. Scales 19 at mid-body..... Ephalophis
15. Scales 25-39 at mid-body..... Hydrelaps
Genera marked with an asterisk (*) are monotypical.
Crocodiles. There are four crocodile species recognized as occurring in the Indonesian Archipelago, namely the gavial Tomistoma schlegelii, freshwater crocodile Crocodylus novaeguinae, the marsh or salt-water crocodile Crocodylus porosus, and Crocodylus siamensis.
Tomistoma schlegelii is found in Sumatra and Kalimantan; their status is unknown. Crocodylus siamensis is believed by many to be locally extinct, however, it may be found in small populations in remote areas of East Kalimantan. Wild populations of C. novaeguineae are restricted to Irian Jaya and Papua New Guinea. C. porosus is the most widely distributed crocodile in the archipelago. Its distribution is patchy, however, and larger populations are now restricted to more remote and inaccessible areas in the eastern parts of the archipelago.
Population densities of all three species of crocodiles have decreased sharply during the past two decades. In many regions of the archipelago, crocodiles were hunted out, and are now locally extinct. The remaining populations are restricted to relatively uninhabited and remote areas of Sumatra, Kalimantan and Irian Jaya. Relatively abundant in former times, all three species are severely threatened throughout their distribution range. Cox et al. (1993) have recently examined a number of freshwater crocodiles in East Kalimantan which closely resembled C siamensis, long extinct on Java and many areas throughout Southeast Asia. The largest populations of C porosus and C. novaeguineae occur in Irian Jaya and are tentatively estimated at 63,000. Efforts are being made to farm and conserve remaining populations.
Box 21.8. Shorebirds in Indonesia.
Y.R. Noor and M. J. Silvius, Asian Wetland Bureau - Indonesia, P.O. Box 254/BOO Bogor 16002, Indonesia.
Marine Birds. Introduction
The Indonesian Archipelago is a country of more than 17,000 islands and many seas that stretch over 5200 km along the equator. Almost 14% of its 7.7 x 106 km2 area is covered by wetlands.
The shallow, muddy coasts of Indonesia are fringed by the world's most extensive mangrove forests, which are regularly flooded with brackish or saline tidal waters. The flood plains, mangroves, and mud flats have the potential to support vast numbers of shorebirds (fig. 21.50). Between September and March, Indonesia's coasts are visited by millions of shorebirds which feed and rest during their migration from the northern latitudes of Asia and Alaska to east and Southeast Asia, Australasia, and islands in the west Pacific. Studies to assess the importance of this region for shorebirds have been few, and most have been carried out only in the last few years.
Sixty-four (64) species of shorebirds have been recorded in Indonesia, of which a few are not regularly observed. There are 14 resident species, but their breeding distribution is not completely known. For the endemics, there is little new information. The Javanese wattled lapwing (Vanellus macropterus) is now considered extinct, with no records since 1937, while the Obi woodcock has not been recorded since 1902.
Shorebird Habitats
Of the Indonesian wetland habitats, some have been identified as suitable habitats for shorebirds, including:
a) Mangroves and mud flats. Indonesian mangroves are among the best developed and diverse in the world, with a particularly rich associated fauna of crustaceans and mollusks. The availability of food makes Indonesian mangroves a preferred habitat for migratory as well as resident waterbirds. These include the endangered spotted greenshank (Tringa guttifer) and the rare Asian dowitcher (Limnodromus semipalmatus), which has its main non-breeding area on the southeast coast of Sumatra.
b) Herbaceous swamps, grass swamps, and savannas. Swamps that are intermittently dry, or shallowly inundated for most of the year, may be of importance to shorebirds.
c) Lakes. The lakes are not known to support large numbers of shorebirds, but may do so during the dry season, when large areas of mud are exposed.

Figure 21.50. Indonesian coastal mud flats are critical habitats for thousands of resident and migrating shorebirds.
Photo by Asian Wetland Bureau.
Shorebird Abundance and Distribution in Indonesia
The vast coastline and wetlands of Indonesia, combined with the difficulty of access, are responsible for the incomplete counts of Indonesian shorebirds. Little is known about the status of shorebirds in the large areas of wetlands which lie outside the sites visited by birdwatchers.
In Indonesia, sixteen sites have been identified as being of international importance for shorebirds. The sites with the largest numbers of shorebirds occur on the north coast of Java and the southeast coast of Sumatra, and these sites account for over 90% of the shorebirds counted (fig. 21.51). Based on the presence of suitable habitats such as mangroves and intertidal mud flats, other coastal areas likely to hold large numbers, but not yet surveyed, include the south and east coasts of Kalimantan, and the unsurveyed sections of the south coast of Irian Jaya.
Of the 64 shorebird species that were recorded and counted, black-tailed Godwit (Limosa limosa), redshank (Tringa totanus), and lesser sandplover (Charadrius mongolus)are the most numerous (fig. 21.52).
Threats to Shorebirds and their Habitats in Indonesia
The two principal threats to shorebirds in Indonesia are habitat destruction or modification, and hunting on a non-sustainable basis. Excessive use of pesticides in rice fields may pose a significant threat, although this remains to be confirmed.
Destruction of Habitat
Habitat degradation caused by loss of mangroves, with a resulting lowering of biological productivity in intertidal mud flats, could affect shorebird populations. It is not known if areas currently supporting shorebirds are at their carrying capacity, or whether a reduction in the density of food caused by lowered biological productivity will automatically lead to reductions in the numbers of shorebirds.
The principal known shorebird habitats in Indonesia, with the exception of those in Java, occur in areas where the human population density is relatively low (i.e., southeast Sumatra, Sulawesi, and Irian Jaya). For this reason, many shorebird sites are still reasonably intact, although a few of them are protected in reserves.
In Java, substantial numbers of shorebirds occur along areas of the north coast, which also support a very high human population density. Despite this, some of Indonesia's most significant shorebird sites still exist on this island. However, a rapidly increasing human population, combined with the rapid pace of development, could lead to a substantial loss of habitat on Java within the next 20 years.
Hunting
Hunting pressures on shorebirds are believed greatest in Java. Data from the north coast of West Java show that at least 300,000 waterbirds are captured every year for food. Hunting pressures in other regions of Indonesia have not been completely evaluated yet, however, the threat is believed to be serious in some areas.

Figure 21.51. Maximum number of shorebirds counted in Indonesia during 1983-1992. Key to locations: SumATRA: 1 - Tanjung Tatuk; 2 - Bekawan Luar, Pulau Burung, P. Ketam, P. Serapang, Tanjung Balai, and Sungai Guntung; 2a - Tanjung Baakung; 3 - Kuala Cemara, Permusiran, Sungai Jambat, S. Sinur Naik, Benlahara, Pangkal Duri, Sungai Benu, Labuan Pering, Tanjung Jabung, Lambar Luar, Sungai Alang-Alang, Kampung Laut; 4 - Banyuasin delta, Sungai Buntol, Pulau Betet, Sungai Betet, P. Birik, S. Nibung, Teluk Gelas; 5 - Rawa Sragi. JAVA: 6 - P. Dua, Sawah Luhur; 7 - Muara Angke, K. Muara, Karang Mulya; 8 - Bungko, Astana; 9 - Segara Anakan; 10 - Solo delta, Brantas delta, Perengan, Semangkan; BALI: 11 - Suwung; LESSER SUNDA ISLANDS: 12 - Panapa, Kawangu, Melolo, Tapil, Warambadi, S. Rende; 13 - Pantai Kupang; KALIMANTAN: 14 - P. Jawa, Muara Ulu, P. Berau, Senipah, P. Bukuan, Tg. Sembilang, P. Layangan; SULAWESI: 15 - Mapie; 16 - Lenteboeng, Ujung Pandang, Maros; 17 - Bulukumba; 18 - Lake Temp, Danau Buaya; 19 - Bajare-Patiro, Ujong Patiro, Palima Bajuwa, Tipulwe, Bawanawatu; 20 - Palopo, Baliase, Malengke, Montalinga, Balise-Wotu, Watulengkau, Teluk Usa; IRIAN JAYA: 21 - Rawa Dembuwuan, Rawa Cumoon; 22 - Wasur National Park.

Figure 21.52. Indonesian shorebird species with population numbers above 10,000.
Pesticides
The uncontrolled use of pesticides in some areas is a severe threat to the ecology of rice-fields and possibly to shorebirds that use this habitat. There is little information available on this in Indonesia, but studies elsewhere have shown that rice fields threatened with pesticides support a lower number of shorebirds.
Marine Mammals
Indonesia remains an important Southeast Asian refuge for a number of large marine mammals, such as the dugong (Dugong dugon). However, very little is actually known about marine mammals in Indonesia, the dugong being the exception, since there is some information on its populations and general distribution patterns. Almost no information exists on the population and distribution of whales and dolphins, even though it is well-known that Indonesian waters are frequently visited by large whales. Whales reported from Indonesia are listed in table 21.23. There are 31 records, of which seven are baleen whales and the rest are dolphins and their relatives (IUCN 1991). Whales are found throughout the archipelago. Some whales have restricted distributions, while others are widely distributed. The warm and highly productive Indonesian seas may be important breeding areas since whales breed and have calves in warmer water. There are sitings of breeding populations in the Molucca Sea, southeast of Halmahera Island (Moosa, pers. obs.). Large pods of humpback whales were spotted off the south coast of North Sulawesi. A large, solitary blue whale was seen just off the Lucipara Islands.
The hunting of dolphins and whales is traditionally prescribed in many parts of the archipelago. The most important (in terms of catch) whaling villages are Lamalera on Lembata Island, Lamakera on Solor Island and Kampung Lewotobi on Flores, all in East Nusa Tenggara. Limited whaling is practiced by some villages on the Kai Islands in the Moluccas. The whaling fleet at Lamalera has been catching up to 30 sperm whales per year for the past three years. It is interesting to point out that the fishermen of Lamalera take only sperm whales and other toothed whales, while the whaling fleet from Lamakera takes the large baleen whales. In addition to whales, the fishing fleets from the two villages take large fish, turtles and dugongs. Whaling in Lamalera is done with traditional boats and harpoons. The whaling fleets on Lembata and Solor support a traditional barter economy that would suffer substantially if the whaling was stopped without providing a viable alternative. Lamalera is easily accessible, yet very little research has been done on the sperm whale populations.
Indonesian waters also support large populations of dolphins, while the deep eastern waters are evidently an important migration pathway for some of the large whales. Occasional mass-strandings of these large marine mammals have been reported.
Table 21.23. List of whales and dolphins reported from Indonesian waters.

Box 21.9. Current status of dugongs in Indonesia.
H.H.de longh, Centre of Environmental Science, Leiden University, Leiden, the Netherlands.
Introduction The dugong (Dugong dugon, Muller 1776) is the only true extant marine herbivorous mammal. Together with the three species of the riverine and euryhaline manatees, they form the Order Sirenia, the sea cows. A fifth species, the Steller's sea cow (Hydrodamalis gigas), which was found in the Bering Sea by Russian sailors in 1741, was driven to extinction in 1768, only 27 years after it was discovered (Norse 1993). The scientific classification of the sirenians (dugongs and manatees) is as follows:

Sirenians evolved from terrestrial herbivores in the early Eocene (38-54 million years ago). Protein analyses have revealed their close relationship to the Proboscidae (elephants) (de Jong et al. 1981). A maximum of 12 genera were present during the Miocene (26-27 million years ago), but at least 10 genera have become extinct (Domming 1978).
The dugong is locally common in many parts of Indonesia, but is threatened in most of its range by habitat destruction, pollution and hunting pressure. Protection and proper management of dugong populations worldwide are needed to ensure the survival of the species, because: 1) it is a unique animal (intrinsic value); 2) it is locally important as a source of protein for many coastal communities; 3) it can be used as a biological indicator for pollution and coastal habitat degradation and destruction; and 4) it is a potential ecotourism attraction.
Distribution and Abundance
At present, dugong habitats cover the shallow tropical and subtropical waters of the Indo-Pacific between longitudes 30° N and 30° S. Extensive seagrass beds form the dugong's primary habitat. Severe hunting pressure in the past has greatly depleted many dugong populations throughout the Indo-Pacific, and especially in Southeast Asia. The remaining groups are scattered over vast areas. Although the coasts of eastern Australia and remote Indonesian islands are inhabited by a fair number of dugongs, they are regarded as a rare and endangered species, and are listed as 'vulnerable' in the IUCN Red Data Book. Figure 21.53 shows the present distribution of dugongs in Indonesian waters.
Scientific information on dugongs in Indonesia is scarce. Samuel Falours, a Dutch artist employed by the United East Indies Company (VOC), described how in 1719 he kept a "sirene" or "mermaid" inside a bathtub on Ambon for four days and seven hours (Peitsch 1992). The dugong originated from Seram. Salm et al. (1982) reported the presence of dugongs in northern Irian Jaya.
Some data are available on dugongs around Kupang Bay (Timor), Arakan Reef (North Sulawesi), the Togian Islands - Teluk Tomini (Central Sulawesi) and other small bays and straits around Sulawesi (Hendrokusumo et al. 1981). In Java, dugongs have been reported from Ujung Kulon National Park and adjacent waters like Miskam Bay, as well as Belambangan on the south coast of East Java. In the Moluccas, dugongs were reported to be fairly numerous around the Aru Islands (Compost 1980).
Figure 21.53. Known distribution of the dugong (Dugong dugon) in Indonesian waters.
Evidence gathered through aerial surveys over small tropical islands with narrow coastal shelves, indicates a dispersed distribution pattern with relatively low numbers of dugongs when compared with Australian continental shelf areas. Aerial surveys in northern Australian continental waters yielded over 150 dugongs per hour (Marsh 1985), while aerial surveys performed in the Lease Islands showed rates of 5-11 dugongs per hour (de longh 1995a), 5.4 and 7.5 dugongs per hour in Palau (Brownell et al. 1981; Rathbun and Ralls 1988), and 9.2 dugongs per hour in the Torres Strait islands (Marsh et al. 1984).
General Biology
Detailed knowledge about the biology, ecology and habitat requirements of dugongs is still lacking. The majority of dugong research has been carried out in Australian waters where many dugongs are accidentally drowned in shark nets set along the coast of north Queensland. Data on carcasses and captive dugongs in Indonesia was collated by de longh et al. (1995g). The study of these carcasses provided information on the anatomy and life history of this species. Due to the remoteness and the inaccessibility of their habitat, field observations on dugongs are mostly fragmentary. An outline on some important aspects of current knowledge is presented below.
Life History and Reproductive Biology
The study of carcasses from Australia, Papua New Guinea and the Lease Islands revealed some information on the life of the dugong and differences between populations. Age has been estimated by counting dentinal growth layer groups in unworn tusks (Marsh 1980). The oldest animals examined were more than 70 years old. At a body length of about 3 m they weigh about 400 kg. The analyses of size measurements of five adult female dugongs from Ambon and Haruku (de longh et al. 1995f) and a live specimen from the Surabaya Zoo indicated that these animals were larger in size, when compared with the data reported by Spain and Heinsohn (1975) for Australian dugongs. Sexual maturity is not reached before the age of 9 or 10 years in both sexes. Sometimes females do not have their first calf until the age of 15 to 17 years. Only one calf is born after a gestation period of 12 to 13 months. Neonates are about 1 to 1.4 m long and weigh 20 to 35 kg (Marsh et al. 1984). After birth, calves remain with their mother for as much as 18 months, suckling as well as grazing. The calving interval is estimated to range from three to seven years. The dugong is a species with a high investment in each offspring and a low reproduction rate. This means that they are very vulnerable to overexploitation.
Adaptations to Aquatic Herbivory
Dugongs are truly aquatic mammals, therefore, unlike seals, they never have to leave the water. Their streamlined bodies have short, rounded fore-flippers and whale-like flukes. The large head has small eyes, no external ear pinnae and the nostrils are located on top of the snout. The light-coloured, almost bare skin is extremely thick and smooth. Sinus-type hairs are scattered over the entire body, and are especially dense around the mouth. The muzzle itself is a complex structure, specialized to grasp plant material. The dentition is well-adapted to grind the food, and the teeth are replaced throughout its life span.
Some studies have focused on the digestive capacities and consumption rates of dugongs. Murray et al. (1977) reported high fibre digestibility in the hindgut of the dugong. In fact, the values were so high compared to those obtained for other herbivores (even foregut fermenters), that Murray et al. (1977) suggested that the values may have been due to problems in the methodology. Lanyon (1992) showed that the GPR (Gut Passage Rate) of the dugong is one of the slowest measured in any mammal. Consumption rates of 26.0 - 47.5 kg wet weight, indicate a very low metabolic rate, similar to manatees, but a comparatively higher consumption rate compared to manatees. Although the manatee and dugong are both characterized by high digestibility coefficients, low gut passage rates, low metabolic rates and susceptibility to cold temperatures, the food available to the manatee is more digestible compared to the main food of the dugong, which are sea-grasses. According to Thayer et al. (1984), a relatively high percentage of the organic matter content of seagrass is fibre. It is therefore expected that the dugong, as part of its Optimal foraging strategy', selects its food for low in vitro digestibility and high carbohydrate and nutrient content.
De longh et al. (1985c) demonstrated patterns of cultivation grazing of dugongs in the Lease Islands in both intertidal and subtidal Halodule-dominated meadows, contradicting Preen (1992), who suggested that in tropical areas dugongs may not practice cultivation grazing, since they do not suffer from nutritional stress.
Food Preference and Feeding Ecology
Even if there is still no general agreement on the dugongs' food, one could summarize the literature on this subject by saying that the dugong feeds on various species of sea-grasses, occasionally feeding mainly on algae. Significant amounts of marine algae have been reported from dugong stomach analyses (Heinsohn and Spain 1974; Marsh et al. 1982; Johnstone and Hudson 1981). These authors suggested that, in most cases, the eating of large amounts of algae was due to the absence of seagrasses as a result of overgrazing or destruction of the seagrass beds by cyclones.
Although dugongs obviously can eat large amounts of algae, they do not appear to be well-adapted for algae digestion (Marsh et al. 1982). The algae fragments in dugong stomachs are reported to be generally larger than those of seagrasses (Lipkin 1975; Marsh et al. 1982). Lipkin (1975) suggested that this may be due to the "more flexible consistency" of the algal material, so that it cannot be chewed adequately. Some early authors refer to the dugong food in general terms only, as being composed of "fungi and submarine algae", "seawater algae", "algae and seagrasses", or "various seaweeds" ([Rattles 1920; Flower and Lydekker 1891; Tate 1947; Throughton 1951; Desmond 1965; Walker 1968]; all in Lipkin 1975).
Although seagrasses are the main food source of the dugong, some animal material has also been found in the stomach contents and faeces (Marsh et al. 1977). It is not clear whether the dugong benefits from consuming animal protein, or if it can be digested at all. Some researchers believe that invertebrates are eaten when seagrass is scarce or of bad quality (Anderson 1989; Preen 1992).
Gohar (1957) claimed that the Red Sea dugong feeds exclusively on the seagrass Diplanthera (Haiodule) uninervis. Heinsohn and Birch (1972) studied the stomach contents of 15 dugongs accidentally caught and drowned in shark nets near Townsville. Six seagrass species, representing four genera, were found after analysis. According to the authors, besides Haiodule uninervis, Cymodocea serrulata was another heavily consumed species.
In line with these findings, Marsh et al. (1982) analyzed the stomach contents of 95 north Queensland dugongs and found that Haiodule was a dominant genus in the food spectrum of dugongs. Of the eight genera of seagrasses known to be available in the study area, six were identified in the stomachs, the exceptions being Syringodium and Thalassodendron. Two to four genera of seagrasses were found in the stomachs of most dugongs. Halodule was found in 95% of the stomachs, followed by Halophila(89%), Cymodocea (61%) and Thalassia (39%). Seagrass rhizomes were present in all stomachs, including that of a neonate calf. The amount of rhizome eaten varied with the type of seagrass, with HaJodule and Halophila ranking highest.
It is notable that in the study by Marsh et al. (1982), Halodule is present in 95% of the stomach contents. This seagrass species, found in the upper subtidal and lower intertidal zones, is in some areas periodically out of reach to the dugongs due to low tides, and therefore, it is remarkable that it is found in such large quantities in dugong stomachs. The importance of "soft" and "sparse" seagrass species such as H. uninervis and H. ovaliswere recently confirmed by Preen (1992), who stated that dugongs tend to prefer pioneer seagrass species with high N levels and low neutral detergent fibre content.
De longh et al. (1995b) found a strong correlation between dugong feeding frequency in a HaJodtyJe-dominated meadow and the total organic C content of below-ground biomass, indicating a dietary selection based on "energy maximization". Field observations also confirm that dugongs in Indonesia and Australia feed regularly on intertidal seagrass beds of Halodule and Cymodocea, exposed during low tide and flooded during high tide (Prince 1984; Brasseur and de Jong 1991). De longh (1995b,c,e,f) confirmed the dietary preference of dugongs in the Lease Islands for the low serai stage species Halodule uninervis and Halophila ovalis. He also found a similar decreasing preferential order as described by Preen (1993) for subtropical Moreton Bay. Dugongs in the tropical Lease Islands appear to have the following preference for seagrasses: Halophila ovalis > Halodule uninervis > Cymodocea rotundata > Cymodocea serrulata > Thalassia hemprichii. Earlier studies indicated that dugongs preferred intertidal H. uninervis meadows (de longh 1995a). Feeding experiments with an adult female dugong in the Surabaya Zoo showed that the animal preferred H. uninervis and Halophila ovalis over Cymodocea rotundata, Syringodium isoetifolium and Thalassia hemprichii (de longh et al. 1995c). Dietary preference in the feeding experiment correlated significantly with in vitro digestibility (IVD), ash-content, Na-content and Ca-content of the seagrass species included in the experiment. The studies mentioned above indicate a consistent dietary preference of dugongs from subtropical areas to tropical areas.
Behaviour
With respect to the feeding biology of the dugong, some of the observations of de longh et al. (1995g) do not correspond with the findings of other authors. Anderson (1981) and Anderson and Birtles (1978), for example, report that dugongs forage at maximum depths of 3 to 4 m in both sublittoral and intertidal areas. Most of the feeding tracks observed in the seagrass beds in Saparua Bay (Saparua Island, the Moluccas) were located at a greater depth (7-9 m). Marsh et al. (1994) and Marsh (1993) reported on the importance of deepwater seagrass meadows to dugongs and found evidence of dugong feeding down to a depth of 40 m. Favoured feeding areas appear to be characterized by relatively sparse seagrass and proximity to deeper water. De longh et al. (1995g) also concluded that there was no indication that dugongs dislike rough water, in contrast to earlier writings (Allen et al. 1976). Even during very rough seas, dugongs would forage. This could be explained either by the importance of the nutritional value of the grasses or the depth of the meadow (7-10 m). Dugongs were spotted feeding between 9:00 and 16:30, suggesting that they do not have regular feeding times.
Anderson and Birtles (1978) state that feeding tracks are only found in Zostera and Halophila, and thought that each trench represented the effort of a single dive. However, de longh et al. (1995g) observed a feeding dugong producing multiple feeding tracks. The dugong's behaviour may be adjusted to minimize effort for maximum energy intake (de longh et al. 1995b). Multiple feeding tracks during one dive support this thesis. During feeding, the dugong displayed no active use of its flippers, in contrast to the observations of Jarman (1966). From our observations it is concluded that the curves in feeding tracks are probably the result of the dugong's haphazard movements.
De longh et al. (1995g) found that the mean submerged time of dugongs is 4.6 min. This is longer than 0.9-2.7 min. (Allen et al. 1976; Anderson 1982), and 0.5-1.5 min. (Anderson and Birtles 1978) in waters of up to 3 m deep. Anderson (1982) concluded that the interval between appearances at the surface varies with locality (habitat), foraging mode, food species, activity and reproductive status. His data suggest that dugongs remain submerged longer in deeper water (i.e., the submerged time correlates with the depth of the bed). From our observations it can be concluded that the dugong breathes more often (up to four times) in rough water than in calm water (once or twice), between dives. In the study of de longh et al. (1995g), the time spent at the surface was a minimum of three seconds for one breath, which agrees with earlier observations (Anderson and Birtles 1978). Maximum time was 1.03 min. for three breaths. The average surface interval between dives was 37 seconds. Territorial lagging behaviour as observed during the present study was only reported by Anderson (1979) for dugongs in Shoalwater and Cleveland Bays in Australia.
Dugongs usually swim slowly (at about the speed of a human diver with flippers), using only their enlarged tail and flukes for locomotion. When disturbed, stronger vertical strokes of the flukes accelerate the movement for a short time. When foraging, dugongs dive to the seagrass beds and graze usually at about 4-5 m depth. They surface for a few seconds to breathe and then dive again (Anderson and Birtles 1978). Calves accompany their mother by swimming almost in body-contact above her back. Only when feeding inshore on small seagrass species in intertidal zones do they produce the characteristic feeding tracks (see 'feeding ecology').
Historical reports suggest the existence of aggregations comprising several thousands of animals in one herd. Where dugongs are still abundant and not hunted (e.g., in Australia), they may still be encountered in large groups, numbering up to several hundred individuals. Solitary individuals are reported from other areas. The social behaviour of this species is still poorly understood. One function of the aggregation in large groups may be predator avoidance (de longh et al. 1995c). This is also supported by the observations of Anderson (1982) in Shark Bay, Western Australia. When he approached a group of dugongs by boat or while diving, some individuals came very close and visually investigated the object, while the rest of the animals remained at a distance of several hundred metres. The eyes of the dugongs are adapted to the low light intensity under water, however, dugongs also raise their head well above the surface and are known to look carefully at approaching objects.
Little information is available on the intra-specific communication of dugongs. Anderson (1982) used sensitive hydrophones close to dugong groups but could not detect specific sounds produced by the dugongs. Aborigines report the existence of whistlers (i.e., the male in charge would warn the group by producing characteristic whistling sounds). In areas where dugongs are severely hunted or otherwise disturbed, it is not possible to approach them so easily.
Conservation of Dugong Populations in Indonesia
Dugongs depend on the production of seagrass ecosystems, and are the only marine mammals which graze intensively on seagrasses. As marine herbivores, dugongs are a potential protein source for coastal communities. However, due to their unknown status and low reproduction rates, it is recommended to maintain and enforce the protected status of this species in Indonesia. In line with the IUCN Action Plan for Dugong Conservation and the Global Plan for the Conservation, Management and Utilization of Marine Mammals, there is an urgent need in Indonesia to conduct more research on dugongs, to prepare population inventories and to establish a national conservation strategy and management plan. Designation of protected areas should receive the highest priority. The main points of action are summarized as follows:
1. Intensified research is urgently needed on population size, distribution, life history, dietary requirements, habitat requirements, behaviour, habitat loss, exploitation and incidental catch of dugong populations in Indonesia.
2. An inventory should be prepared of current conservation measures, such as legal protection, traditional conservation, existing marine parks covering dugong habitats, and potential marine park areas covering important dugong habitats.
3.A national conservation strategy for dugongs should be prepared, starting with a regional and nationwide resource inventory, including the establishment of a database on distribution and abundance of dugong populations, definition of the goals for dugong conservation, habitat surveys, and assessment of the status of dugong populations.
4. Based on the conservation strategy, a dugong management plan should be established, as part of an integrated coastal zone management plan. An important component of this plan should be the establishment of protected areas, covering important dugong habitats. Complementary zoning and appropriate land use on the adjacent coast is needed because of the interdependence between land and sea.
5. The success of a dugong conservation strategy will depend to a large extent on the concern and cooperation of the public. A programme of information dissemination needs to be undertaken in Indonesia, both though government programmes and with the assistance of NGOs.
References
Allen, J. F., Budiarso, M. M., Sumitro, D. and Hammond, D. (1976). Some observations on the biology of the dugong (Dugong dugon) from the waters of South Sulawesi. Aquat. Mamm. 4: 33-48.
Anderson, P. K. (1979). Dugong Behaviour: On being a marine mammal grazer. The Biologist 61(4): 113-144.
Anderson, P. K. (1981). River Cows and Sea Pigs: A proposal for revision of Sirenian popular names. Sirenews 16: 3-4.
Anderson, P. K. (1982). Studies of dugongs at Shark Bay, Western Australian. II Surface and subsurface observations. Aust. Wildl. Res. 9: 85-99.
Anderson, P. K. (1989). Deliberate foraging on macro-invertebrates by the dugong. Natl. Geogr. Res. 5(1): 4-6.
Anderson, P. K. and Birtles, A. (1978). Behaviour and Ecology of the Dugong, Dugong dugon (Sirenia), Observations in Shoalwater and Cleveland Bays, Queensland, Aust. Wildl. Res. 5: 1-23.
Brasseur, S. and de Jong, G. (1991). Survey of the Dugong in the Moluccas and the recolonization of their feeding tracks. Report AID Environment, p. 89.
Brownell, R. L. Jr., Anderson, P. K., Owen, R. P. and Ralls, K. (1981). The status of dugongs at Palan, an isolated island group. In: Marsh, H.(ed.), The Dugong. Proceedings of a seminar/workshop held at James Cook University of north Queensland, 8-13 May 1979. Townsville, James Cook University, pp. 11-23.
Compost, A. (1980). Aru Islands, Indonesia. Dugong and turtle survey. Project 1734. WWF Monthly Report, April 1980.
de longh, H. H. (1995a). Aerial Survey of the Dugong (Dugong dugon, Miilier 1776) in Coastal Waters of the Lease Islands, East Indonesia. Australian Journal of Marine and Freshwater Research (in press).
de longh, H. H., Wenno B. J. and Metis, E. (1995b). Seagrass distribution and seasonal biomass changes in relation to dugong grazing in the Moluccas, East Indonesia. Aquatic Botany (in press).
de longh, H. H., Hein, L. and Burger, M. (1995c). Modelling of dugong feeding pressure by snorkeling surveys of seagrass meadows in Haruku Strait, East Indonesia. Marine Ecology (in press).
de longh, H. H., and Langeveld, P. (1995d). Cultivation Grazing of Dugongs in Haruku Strait, East Indonesia (unpublished).
de longh, H. H., van Langeveld, P., Van der Wal, M. and Norimama, D. (1995e). Movements and home ranges of dugongs around the Lease Islands, East Indonesia. Journal of Applied Ecology (in press).
de longh, H. H., and Bauer, H. (1995f). Dietary preference of a captive held dugong in Surabaya Zoo, Indonesia, Ecology (in press).
de longh, H. H., van Bierhuizen, B., and Waisfisz, Q. (1995g). Observations on size and behaviour of the dugong (Dugong dugon, Muller 1776) from the waters of the Lease Islands, East Indonesia, Biological Conservation (in press).
De Jong, W. W., Zweers, A. and Goodman, M. (1981). Relationship of aardvark to elephants, hyrexes and sea cows from a-crystallin sequences. Nature: 292: 538-581.
Doming, D. P. (1978). Sirenia. In: Maglio, V. J. and Cooke, H. B. S. (eds.), Evolution of African Mammals. Harvard Univ. Press, Cambridge, pp. 573-581.
Gohar, H. A. F. (1957). The Red Sea dugong. Publ. Mar. Biol. St. Ghardaga 9: 3-49.
Heinsohn, G. E. and Birch, W. R. (1972). Food and feeding habits of the dugong, Dugong dugong (Erxleben), in northern Queensland, Australia. Mammalia 36: 414-422
Heinsohn, G. E. and Spain, A. V. (1974). Effects of tropical cyclone on littoral and sub-littoral biotic communities and on a population of dugongs [Dugong dugon (Muller)]. Biol. Conserv. 6: 143-152.
Hendrokusomo, S., Sumitro, D. and Tas'an (1981). The distribution of the dugong in Indonesian waters, In: Marsh, H. (ed.), The Dugong. Proceedings of a Seminar/Workshop held at James Cook University 8-13 May 1979. James Cook University, Townsville, 240 pp.
Jarman, P. J. (1966). The status of the dugong (Dugong dugon Muller). East Afr. WildlifeJ. 4: 82-88.
Johnstone, J. M. and Hudson, B. E. T. (1981). The dugong diet: mouth sample analysis. Bull. Mar. Sci. 31:681-690.
Kingdon, J. (1971). East African mammals, an atlas of evolution in Africa. A Rad. Press,
London, New York, 446 pp. Lipkin, Y. (1975). Food of the Red Sea Dugong (Mammalia: Sirenia) from Sinai. Israel Journal of Zool. 24: 81-98. Lanyon,
J. (1992). The nutritional ecology of the dugong Dugong dugon in tropical N. Queensland. Department of Ecology and Evolutionary Biology, Monash University, Victoria, Australia, 326 p.
Marsh, H. (1980). Age determination of the dugong [Dugong dugon (Muller)] in northern Australia and its biological implications. Rep. Int. Whal. Comm. Spec. Issue 3: 181-201.
Marsh, H. (1985). Results of the aerial survey for dugongs conducted in the Cape Bedford to Cape Melville area in November (1984). Report to the Great Barrier Reef Marine Park Authority. James Cook University, Townsville, Queensland, Australia.
Marsh, H., Channels, P. W. and Heinsohn, G. E. (1982). Analysis of stomach contents of dugongs from Queensland, Australia. Wildlife Res. 9: 55-67.
Marsh, H., Heinsohn, G. E. and Marsh, L. M. (1984). Breeding cycle, life history and population dynamics of the dugong, Dugong dugon. Aust. J. Zool. 32: 767-788.
Marsh, H., Heinsohn, G. E. and Spain, A. V. (1977). The stomach and duodenal diventricula of the dugong (Dugong dugon). Functional Anatomy of Marine Mammals, part 3, edited by Harrison, R. J. Academic Press; London, NY, San Francisco.
Marsh, H., Barker-Hudson, B. E. T., Heinsohn, G. E. and Kinbag, F. (1984). Status of the dugong in the Torres Strait area: results of an aerial survey in the perspective of information on dugong life history and current catch levels. Report to the Australian National Parks and Wildlife Service.
Murray, R. M., Marsh, H., Heinsohn, G. E. and Spain, A. V. (1977). The role of the midgut caecum and large intestine in the digestion of sea grasses by the dugong (Mammalia: Sirenia). Comp. Biochem. Physiol. A 56: 7-10.
Peitsch, T. W. (1991). Samuel Falloura and Lis "Sirenne" from the province of Ambon. Archives of Natural History 18(1): 1 -25.
Preen, A. (in press). Interactions between dugongs and seagrasses in sub-tropical environments. Thesis submitted for the degree of Doctor in Philosophy, Dept. of Zoology, James Cook University of north Queensland, Australia, 265 pp.
Prince, R. I. T. (1984). Dugong in northern waters of Western Australia 1984. Technical Report No. 7, Department of Conservation and Land Management, W.A.
Rathbun, G. B. and Ralls, K. (1988). Status of dugongs in waters around Palau. Marine Mammal Science 4(3): 270-265.
Salm, R. V. and Clark, J. R. (1984). Marine and Coastal Protected Areas: A Guide for Planners and Managers. IUCN Workshop World Congress on National Parks, Bali Indonesia, pp. 301.
Salm, R. V., Petocz, R. C. and Soeharlono, D. T. (1982). Survey of coastal areas in Irian Jaya (1982). Directorate of Nature Conservation, Directorate General of Forestry, Republic of Indonesia (unpublished report).
Spain, A. V. and Heinsoh, G. E. (1975). Size and weight allometry in a north Queensland population of Dugong dugon (Muller) (Mammalia: Sirenia). Aust. J. Zool. 23: 159-168.
Thayer, G. W., Bjomdal, K. A., Ogden, J. C, Willimas, S. L and Zieman, J. C. (1984). Role of large herbivores in seagrass communities. Estuaries 7: 351-376.