10
When 10 000 wildebeest begin a process of migration and they move together, there is no guarantee that any individual knows another single other individual in the crowd. There may be dyads of mother and offspring but, on the whole, a high degree of anonymity can be maintained. A high degree of anonymity can also be maintained in sexual reproduction because individuals might meet briefly to perform the sexual act and then never meet again. Crowds and sexual partners thus do not guarantee company. Anonymity requires very little communication apart from a few ritualised acts that are usually sufficient to trigger a response in another or in an entire flock. An entire flock may rise and take flight at the instigation of one taking to the air. Many of the polygynous males that display or sing to attract a female will not know, and probably would not even recognise again, the females with whom they copulated because they may not even be likely to see them again and have no part in raising young.
The ability for collective action from insects, to birds, to ungulates and primates has become a hot topic in ethology (Giardina 2008) and debates have now moved into the cognitive area, but these pose different issues and questions and may require mathematical modelling to determine the information flow in ant colonies or even flocks of birds.
One of the puzzling phenomena is that some classes or some orders of animals have developed ways of staying together and of investing a good deal of time and care into maintaining a bond. That is as true of some insect groups as it is of fishes, birds and mammals. Of all classes of animals, perhaps birds have pushed the experiment of losing anonymity in favour of commitment and mutual support to the greatest extent. A permanently bonded pair of galahs may be together for 60 or more years, sulphur-crested cockatoos even longer. There is no other class of animals that has such a high incidence of pair bonding and joint parenting as do birds, and, in Australia, this is often coupled with extraordinarily long-term commitments.
Add to this the high incidence of cooperative breeding in Australia and it can be seen that dyads and larger groups deserve much more research attention in terms of the regulatory mechanisms of such close living than they have received so far.
One of the inevitable consequences of losing anonymity is that ways have to be developed in which individuals (a) have to recognise each other reliably every time and (b) somehow integrate their activities with each other. In these contexts, then, communication may become very important and the question is how this is achieved. The social brain hypothesis, as mentioned before, has long argued that evolution of such lifestyles requires complex cognition. In other words, regulatory mechanisms need to be in place to avoid too much conflict, too many distractions, misunderstandings or even failure of group cohesion. Some such mechanisms may be simple, and even adaptive, while others have been shown as cognitively well advanced.
General features of vocalisations
One of the hallmarks of communication in birds is vocalisation. Apart from humans, no other class of animals has developed such a plethora of vocal signals as have birds. These have been a rich source of study for over 100 years. Now technology has aided any inquiry by allowing simple transfer of sounds onto sound-converting programs showing up as two-dimensional sonograms that can be analysed for all the elements that make up sound. Since the 1990s, such programs have become very sophisticated and detailed and they can even perform the relevant calculations. Here, technological advance has given us a major tool and a window into the communication of birds. Over the last decades, meaning and purpose have been discovered in bird sounds that were once just appreciated for their musical or interesting sounds and some of these will be highlighted in this chapter.
Frequencies of bird vocalisations commonly range within comfortable human auditory range, between 2 and 10 kHz, and are therefore easy to record and measure. Only a few bird species are known to produce infrasounds, such as the pigeon (sound levels down to 0.5 Hz). A few, such as oilbirds and cave swiftlets, can even produce vocalisations, or at least perceive sounds, in the ultrasonic range between approximately 20 kHz and 100 kHz (Konishi and Knudsen 1979; Brinkløv et al. 2013).
Sending a vocal message can take many different forms. For birds (and many insects) with a high investment in communication by acoustic means, effective sending of messages can be quite difficult. It can be impaired by auditory saturation, for instance, when sounds are impossible to transmit over a long distance because of background noise (such as wind, rain, or noise from insects and frogs) or aggregate noise produced by the same species living communally in a colony, in large family groups or in a bachelor flock. The receiver of a vocal signal must, therefore, have capabilities of extracting the relevant information from random background noise and must have the capacity to detect information-carrying signals of an intensity that may be below that of the background noise. A message, even at its most basic level, also reveals the species identity of the sender.
Repertoire size may vary widely between species. The champion with the greatest repertoire in Australia is likely to be the Australian magpie with roughly 800 syllables identified (Farabaugh et al. 1988) to make up an endless variety of songs (Kaplan 2006). Worldwide, it may be the brown thrasher with between 1500 and 1900 song types (Catchpole and Slater 2008).
Song size, by itself, is not an indication of an increase in meaning. The functions that have been established specifically for birdsong can be summarised, with some exceptions, as territorial defence and sexual attraction. But then there is a whole range of calls that can be very brief and these can have very different functions. They can signal about food, can express anxiety or alarm, rivalry, attention, defence, flying away (follow me) and similar short instructions. None of these may be specific to the sex of a bird. Male and females alike will utter calls in relation to predators and a host of other situations, but this depends on the context. At its most basic level, a vocal message would reveal the identity of the species because birds have species-specific vocalisations, and these may consist of syntax and the semantics of a message. The syntax refers to the structure of the song or call. Semantics refers to the content or meaning of the message. Both can be intertwined. ‘Vocabulary’ means the same as the semantics (i.e. the actual meaning of the calls).
The multiplicity of functions of bird vocalisations, while now well understood, still tells us relatively little about their cognitive dimensions. The question that has interested researchers is whether vocalisations (other than crystallised song) are mere expressions of affect (emotions) or are produced intentionally by the sender and whether the receiver takes note of the call whether made intentionally or not and can decipher the context in which it is made.
Duetting
The simplest and most common form of daily reconfirmation of bonding is duetting, widespread in the tropics and in Australia. It seems to occur more frequently in pairs with a prolonged monogamous bond. Of all forms of communication, duetting may have least to offer in terms of investigations into cognition but it is an important step in bond maintenance and requires some learning. One bird of a pair initiates a call and the other one answers, usually involving sequential calling rather than, as used in human song, singing together.
Duets may overlap but usually the calls of two birds follow each other so closely and so precisely that they may sound like the vocalisations of one bird (Fig. 10.1). As Hall and Magrath (2007) found, among other things, pairs of magpie-larks are more likely to sing such well-coordinated and precise duets when well established, while those of newly mated birds tend to overlap.

Fig. 10.1. Spectrogram of a magpie-lark duet. Each caller (marked ‘a’ and ‘b’, respectively) emits two sounds: (a) the male initiates the duet and the only noticeable difference between the pair is that the male is a fraction of a second faster in replying than the female (b). (Author’s own recordings.)

Fig. 10.2. Pheasant coucals are unusual birds. They move on the ground and under the shrub in an almost reptilian fashion, they are very poor flyers and usually have to hop along branches to reach the top and they rarely do so (usually only when they need to dry their plumage after heavy rain or when duetting). Pheasant coucals sometimes even raise two broods a season, pair bond and duet, with the male being the major care-giver for the brood. This is a rare photo of a male pheasant coucal sitting on the very top of a large mango tree (taken by author in Coffs Harbour). The highly exposed bird then commenced a loud duet long distance across the valley.
As can be seen in Fig. 10.1 one bird initiates a call and the other one answers. From this Brumm and Slater (2007) concluded that the temporal precision of a duet could reveal to others that the duetting pair is an experienced team and thus attest to the quality of their coalition and their ability to cooperate (which in turn could make the pair more intimidating to a potential intruder).
One of the duetting birds in Australia, the pheasant coucal, duets by overlapping and does so over substantial distances. The two duet type calls known also carry for at least a kilometre (Maurer et al. 2008). It is, as said previously, one of the few cuckoo species worldwide that raises its own young and is noted also for its male-biased parental care and reversed sexual size dimorphism (Taplin and Beurteaux 1992; Maurer et al. 2011), as is the case in most Australian birds of prey. Pheasant coucals usually scuttle along the ground but when duetting, they usually seek a high vantage point (Fig. 10.2).
The functions of duetting are generally well known (Table 10.1). A review on duetting worldwide mentions only seven Australian species in its tally, however: these are the pheasant coucal (Maurer et al. 2008), loriine parrots (Serpell 1981), purple-crowned fairy-wren (Hall and Peters 2008), Australian magpie (Brown and Farabaugh 1991), Australian magpie-lark (Tingay 1974; Hall 2000; Hall and Magrath 2000; Mulder et al. 2003; Rogers AC et al. 2004; Hall and Magrath 2007); eastern whipbirds (Watson 1969; Rogers 2005; Mennill and Rogers 2006; Rogers AC et al. 2006, 2007) and the zebra finch (Elie et al. 2010).
Table 10.1. Duetting: characteristics of participants (based on 48–59 species), after Dahlin and Benedict 2014.
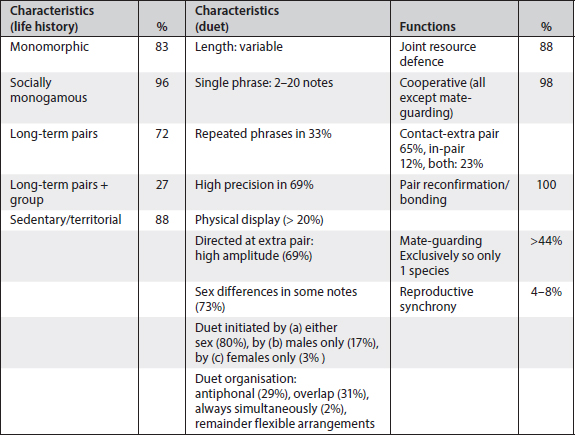
There are probably many more species that duet than just those seven. Informally, we know that the black-faced cuckoo-shrike, powerful owl, tawny frogmouth (Kaplan 2007a) and the southern boobook owl duet, although there exists some controversy about the interpretation of the calls (Farabaugh 1982; Olsen and Trost 2007).
Studies by Charles Blaich and colleagues found that pair-bonded zebra finches engage in contact-call duets far more frequently and in a non-random fashion than unpaired finches (Blaich et al. 1996). In all, Dahlin and Benedict (2014) concluded: ‘All evidence suggests that duets have evolved to advance the interests of a joint partnership. Even the mate-guarding hypothesis rests on the premise that duetting partners cooperate to raise young’.
Hall and Magrath (2007) have made an even more important argument. They have shown that the temporal coordination in the duet indicates the quality of a coalition. This is an important point to make because it suggests a learned part of the alliance. Hence, even in pair bonds, the quality and consistency in cooperative behaviour become a lynchpin for survival.
Alarm calls, mobbing in territorial and nest defence
Territorial defence by vocalisation is generally regarded as a more efficient way of communicating than physical confrontation. Less energy is expended in the process and injuries may also be minimised. Many bird species first issue warning calls to an invader but direct flight at the invading individual may follow if the warning was not sufficient to deter an invader.
Of all call groups, alarm calls have been the most widely studied in animals (Hanson and Coss 2001a,b; Blumstein et al. 2004; Hollén and Manser 2006), in birds in general (Marler 1955; Evans et al. 1993; Davies et al. 2006; Hollén and Radford 2009) and in Australian native birds as well (Jurisevic and Sanderson 1994 a,b; Kaplan 2008a; Kaplan et al. 2009; Fallow et al. 2013). Alarm calls are now recognised generally as reliable signals that, in some confirmed cases, are thought or known to be referential (more of this below).
The loudness of a vocalisation (amplitude) can make a substantial difference to a message. Many bird species, as Richard Andrew (1956) found, have loud–faint pairs of song display. The loud vocalisation may be for territorial display and can mean that the caller would attack if the territory boarders were infringed. For instance, Australian magpies have a vocalisation display that is uttered only when ready to attack. A fainter call signalled to mates and offspring may indicate that the communicator is ready to interact but not attack.
Marler (1955) noted many years ago that warning calls about aerial predators have similar acoustic qualities among very different species of birds and are delivered with approximately similar intensity and at about the same pitch of 7 kHz: an ideal frequency to make localisation difficult. Usually both ears (binaural) assess and compare crucial elements of a sound such as phase, intensity and time difference, thereby decoding message and location of the sender. Phase differences (referring to the timing of a sound wave reaching first one ear and then the other) can be detected more effectively at low frequencies. At higher frequencies wavelength decreases, rendering phase difference more difficult to detect hence the origin of the sound more ambiguous.
In fact, depending on the distance between the listener’s ears (i.e. the size of the listener’s head), there are frequencies of sound that make it impossible to assess location and direction of the sound source altogether because of the difference in timing of arrival of the sound wave at each ear. Were this to be used as the frequency of the alarm call, the predator would be completely unable to use timing to locate the prey.
Identifying the location of the sound source is furthermore aided by the so-called sound-shadow intensity effect. If, for example, the sound source is to the listener’s right, the left ear will be in the ‘sound shadow’ of its head. An intensity difference thereby occurs between ears and this can help to establish the direction from which the sound emanates. If a caller wants to avoid detection, a call ought to be pitched at a frequency that can make phase difference ambivalent and minimise the sound-intensity effect. Marler showed that a call of about 7 kHz does exactly that. This means that certain sets of alarm calls may become common and recognisable to many species because of their physical properties. Discovery of such rules of communication also make it more understandable why communication between very different species is possible. For instance, alarm calls issued by noisy miners often result in magpies rushing to the scene ready to intervene, and vice versa. Birds of prey may be mobbed jointly in acts of mutualism and shared interests.
Marler’s (1955) important finding of the merits of alarm calls at a frequency of 7 kHz (and therefore remaining undetected by the bird of prey) does not hold for magpies. Their frequency is somewhere between 3–6 kHz, depending on urgency and whether issued in the air or from a perched position. Yet magpies are the police of the bush and almost all species respond immediately to their calls. This does not invalidate Marler’s findings. The difference is that magpies, like crows in North America, belong to the group of birds that actively defend their territory and will fearlessly face a bird of prey substantially larger than themselves and mob it whether on the ground or in the air in order to move the predator on and out of their territory (Fig. 10.3).
Mobbing behaviour
Mobbing behaviour, often following alarm calls, may be regarded as one of the most pronounced declarations of cooperation among Australian birds. It happens most often at breeding times near a nest site and involves one partner while the other is brooding.
In cooperatively breeding species, mobbing becomes a highly coordinated event. We tested this in the field by placing taxidermic models of various local and known predators in the field. First on the scene were usually the magpies, but if the problem persisted and the territory was shared with noisy miners, cooperation extended to cross-species support. Goshawks and little eagles were particularly vigorously mobbed with good reason: both are bird hunters and a risk to juveniles and adults alike. Mobbing behaviour is especially widespread among cooperative bird species in Australia. It signals coordination of attack and risk taking for the sake of the pair or the group. In magpies, mobbing attacks are executed in relays and with great precision. When the predator has moved on, the pair or group flies to a high position and engages in a bout of joint victory carolling, further cementing their cohesion (Kaplan 2008). More than that, magpies recognise specific qualities about predators and may choose different strategies of attack depending on the agility and size of the predator, its state of health and general demeanour. As described in detail elsewhere, magpies are clever strategists and these tactics are known and practised by all adult members of the group (Koboroff et al. 2013).
Indeed, there are many Australian songbirds, even quite small ones, that will give an alarm call and approach a predator or intruder. Masked lapwings, very familiar defenders of nest sites, are noted for their fearless attack of pedestrians if deemed to be too close to a nest site. Even the small fantails (rufous and grey) will approach humans, gesture and vocalise incessantly flashing their tails, to make their displeasure known.
Mobbing is not just confined to attacking predators but, as has been found more recently, may also include brood parasites. In an important paper by Langmore and colleagues, it was shown convincingly that speckled warblers and superb fairy-wrens had learned to recognise the bronze cuckoo and mob this enemy jointly before the cuckoo had the opportunity to deposit an egg in the nest of the intended hosts (Langmore et al. 2012). In an interview, Langmore noted that ‘If there are four or more individuals in the group they almost never get parasitised by a cuckoo whereas the small groups of two or three individuals are much more likely to get parasitised. Such mobbing then protects the group and ensures that its own offspring have a chance to survive, an outcome that provides a very strong selective force for the maintenance of cooperative breeding’ (Salleh 2013).

Fig. 10.3. Aerial mobbing behaviour. (Top) An Australian magpie pursuing a brown goshawk. (Bottom) Two magpies mobbing a sea eagle. In both cases, the raptor is either more agile or more powerful than the magpie. The pursuit of the goshawk can only include tearing at the tail feathers because the goshawk can flip around mid-air and also strike out sideways with its talons. The sea eagle can roll around mid-air and a talon is large enough to close around a magpie. The one advantage is that sea eagles are slow flyers and magpies can take the time to swoop. While carrying out this dangerous manoeuvre, they do not vocalise at all. They usually succeed in escorting a raptor out of their own territory on the basis of precision flying and perfect coordination.
A paper from Spain (Canestrari et al. 2014), however, found a new type of interaction between a cuckoo (the great spotted cuckoo) and the carrion crow, by showing that more crow offspring survived when they allowed themselves to be parasitised. The researchers identified a malodorous cloacal secretion that the cuckoo chicks void. They then analysed the secretion as consisting of acids, indoles, phenols and several sulphur-containing compounds that are known to repel mammals and birds. They then recorded the survival rate with and without the secretion and also tested the secretion on cats, by smearing some favourite food with the cloacal mix and other uncontaminated samples. The cats never touched the meat smeared with the compound. They found that it kept predators away and achieved higher survival rate of their own chicks (Canestrari et al. 2014). Mutualism is not cooperation but it is a variant that the group of crows must have discovered because, according to the findings, they stopped attacking the cuckoo and literally allowed themselves to be parasitised. It is not clear whether any species of cuckoo in Australia has developed any similar defence. In most host species in Australia though, it is a definite disadvantage to be parasitised, and hence mobbing may be the only frontline defence against a cuckoo.
Perhaps the majority of short signals addressed to group or partners have to do with defence and are expressing warning or alarm or, at times, also distress. Other short group communications intended for a group or partner may involve announcing the discovery of a new food source.
Intentionality
For observers to prove that any act was performed intentionally in birds has been difficult because the intention itself is prior to any act and only the act itself is visible to us. Hence, any statement in this regard might well appear post hoc. On the other hand, it is difficult to think that alarm calls could ever not be ‘intentional’ in the broadest sense and if some species have more than one alarm call, then it raises further questions. In magpies, Kaplan (2005) discovered 28 variations of alarm calls and such enormous variety does require explanation.
However, Evans (1997) has partly overcome this problem by suggesting that any claim of intentionality must be supported by evidence that the bird had an audience. If an act of communication is intentional, then the bird should be silent even if exposed to the same stimulus if there is no one to whom the message can be broadcast. He tested this by placing a cockerel on his own and exposing it to a bird of prey overhead. While the bird crouched and showed signs of fear, he made no calls. When the same experiment was repeated but, this time, with hens nearby, he issued a clear set of alarms and the hens (that were unable to see the bird of prey) responded also in the same manner of crouching.
It is now known, and still being discovered in more bird species, that birds have what is called ‘external referents’ resulting in referential signals. This means that they refer to objects outside themselves with specific meaning and communicate this to others.
Only two Australian species have so far been tested for referential signalling and these are the budgerigar and the Australian magpie. A recent study by a Dutch team (Van Heijingen et al. 2009) found that budgerigars can discriminate syntactic structures by using their own warble vocalisation in the experiments. We examined the alarm calls of magpies (Kaplan et al. 2009) and found evidence of referential signals specifically for raptors. Furthermore, we found evidence that such signals remain stable and do not lose their impact even in an exported group of magpies (to Fiji) that had no further contact with magpies from Australia but maintained these referential signals even over several generations (Kaplan and Rogers 2013).
The quality and complexity of bird vocalisations has also raised the issue of whether some bird species may be capable of approximating aspects of language (syntax and aspects similar to grammar rules). As already mentioned, Alex, the African grey parrot, understands commands and concepts and can respond to them (Pepperberg 2006 a,b,c). Japanese quail have been shown to be able to learn phonetic categories (Kluender et al. 1987). An even earlier study (Heinz et al. 1981) showed that blackbirds and feral pigeons, both introduced species in Australia, discriminate between vowels. Budgerigars can do such tasks with ease (Tu et al. 2011a,b), challenging the view that speech sound classifications were a uniquely human invention that led to the evolution of language. It indicates the beginnings of a lexicon, of sounds being invested with specific and readily understood and unambiguous meaning, and is generally regarded as a cognitively advanced stage of using vocal signals.
Intentional communication in human society in anything that requires decision making quite often requires negotiation. Negotiation may also be imaginable in birds in which roster systems have to be in place, even if only in rudimentary form – whether for sentinels, for organising the supervision of youngsters in a crèche or for helping in the feeding of offspring. Moreover, negotiations about rosters tend to largely concern future events. Indeed, there has been a study to investigate negotiations in African babblers. The study was conducted observing a wild population of cooperative pied babblers at the Kuruman River Reserve in South Africa (Bell et al. 2010). They found in fact that individuals made anticipatory adjustments to personal investment based on information about the likely future contributions of collaborators. These are very exciting findings indeed, showing that this particular species does not just vocalise intentionally (for which an audience is needed) for an event in the present, but also suggests that this species has a sense of a time scale of things that have not yet happened. One could say that the collaborators ‘discusses’ future plans of action and base their actions on what the intentions of others revealed. To my knowledge, there have been no studies of Australian cooperative species that have tested vocal negotiations of divisions of labour and levels of contribution to a necessary and shared task.
Clearly, this is cognitively complex vocal behaviour and this particular study has opened the doors to testing the possibility in the many cooperative species in Australia – whether or not this is adaptive (i.e. already part of the innate repertoire of the species) or cultural via the avenue of memory, learning and application of cognitive skills is not clear.
Deceptions and cheats
The idea of deception is an interesting category when it is not adaptive mimicry as described in Chapter 7. There have been many anecdotal reports of deception in primates (Byrne and Whiten 1988), but few in birds. The question is in what way deception fits into a topic such as cognition, communication and into the social brain hypothesis. The reason why acts of deception have rarely been reported is that deception works by surprise. To generate the same element of surprise and success again and again is generally questionable. Deception certainly fits into a chapter on communicating emotions. So far, only so-called honest signals have been reported. Except for the example of feigned anger in a juvenile magpie (Fig. 9.12), there are no published examples of feigning anger, frustration or threats.
Dishonest signals can be vocal and the majority of attempts at deception may indeed be vocal in birds. In such cases, a bird has learned to give a false signal with the express purpose of benefitting the caller rather than the recipients of the call and achieving a goal that might otherwise not be achieved (i.e. it requires others to be fooled into actions that they would not otherwise take). One simple means is to sound an alarm call by mimicking that of another species and make all those birds clear the area, allowing the caller to feed on the spoils (Munn 1986; Møller 1988; Flower 2011) or proceed to give a signal that promises one thing and, for the caller, achieves quite another aim. For instance, in domestic chickens, males often call their females to a new site by giving a food call and such a call then results in the females finding a new food source. Hence, most of the time the signal is honest and he delivers on the promise of food. Occasionally, however, the male gives a food call and then uses the proximity of the females to copulate with them (Gyger and Marler 1988). As food calls may also contain information about food quality, the signal may be quite specific but the social context can vary substantially. Smith (1977) considered this issue at length, and concluded that because food calls are given in a wide range of situations not necessarily related to the presence of food as a referent, the call may simply indicate readiness to engage in social interactions. Indeed, Gyger and Marler (1988) retested the behaviour of the cockerels and found that ambiguities remained because the occurrence of the incorrect calls (without food) simply occurred too often, although deceptive use was not ruled out (Gyger and Marler 1988).
‘Crying wolf’, i.e. issuing purposely false alarm calls (Munn 1986), has been reported in several birds: in one case (great tits) to acquire resources (Møller 1988) or, as in some species of drongo, to flush out insects following the commotion of a flock taking to the air (Satischandra et al. 2010). The fork-tailed drongo of South America was recently discovered not just to use deceptive mimicked alarm calls to steal food (Flower 2011), but also to vary the types of alarm calls issued. As said above, trickery is difficult to uphold because the same trick may not work a second time when individuals repeatedly interact and will likely discover they have been deceived and thus not fall for the same trick a second time. Fork-tailed drongos have learned to circumvent this problem by varying the alarm calling they issue to others (Flower et al. 2014). This is termed tactical deception and has been judged as evidence of mental attribution (i.e. knowing what another will do in advance), and thus akin to the most abstract and complex form of cognition: that of theory of mind (Mitchell and Thompson 1986).
Another of the more widespread instances of behavioural deception has been found in cooperative species, specifically concerning helpers that seem to pretend to feed offspring and return to the nest either without food or, if not watched, consume the morsel of food taken to the nest for the nestlings themselves. This has been shown to occur in young white-winged choughs assigned to help feed offspring. They have been shown to feign the feeding motion but then swallow the morsel themselves (Boland et al. 1997b). The same behaviour has also been studied in another Australian cooperative species: the bell miner (McDonald et al. 2007; 2008a,b) and in the cooperatively breeding Spanish carrion crow (Canestrari et al. 2004, 2010), but not been confirmed in the Australian chestnut-crowned babbler (Browning et al. 2012; Young et al. 2012).
Deception in sexual indiscretions seems a little far-fetched, but there are known cases in bonded pair or hierarchical group systems, such as in superb blue-wrens and in magpies, in which cheating males seem to place a good deal of emphasis on secrecy for extra-marital matings. This is now documented in many species. In birds, it is now common to speak of social monogamy and pair bonding, while it is well known from various DNA studies that the social male parent may not be the biological parent. These have been studied most intensively in various species of fairy-wrens (Mulder and Langmore 1993). Many argue that these are adaptations.
More relevantly here, the evidence of behavioural deception, played out specifically in cooperative species would warrant much further investigation. The studies so far undertaken in cognitive ornithology suggest indeed that cheating, or being purposely deceptive, within a group requires complex cognitive skills. As in caching behaviour, outlined in Chapter 3, the apprehension of a possible witness and the knowledge that bad could come from being detected involves knowing the state of mind of another and of consequences that are still in the future and not immediate, and are hence cognitively complex.
Signals for enticement and trickery – the luring game
On an individual level, Robert F. Leslie described an extraordinary case of deception based on interspecies communication between two birds – a parakeet and a blue jay – playing a luring game. The visiting parakeet, perched at the outside of the jay’s cage, seemingly hungry, indicated by eye position and other cues that it wanted the chopped spinach inside the cage. The blue jay moved the chopped spinach close to the edge of its cage, but on the inside, and when the parakeet reached for the spinach, the blue jay delivered a sharp attack on the parakeet’s head (Leslie 1985). The cognitive aspects of this interaction are staggering: it would require the jay to have understood the intention of the parakeet from its body movements and eye movements alone and indeed, the jay’s actions would have needed to be intentional, including a plan to trap the parakeet in order to attack it (an ambush). Perhaps the jay’s behaviour was intentionally malicious and such behaviour requires intelligence.
Probably the most elaborate plot of deliberate deceit is to kidnap youngsters from another group. While some bird species, such as kookaburras, are sometimes willing to adopt orphaned juveniles, it is a very rare behaviour in the animal kingdom to try and kidnap youngsters from another individual, pair or group, except in baboons and some other primates. Primates grab youngsters from females but how can a juvenile bird be made to join the kidnapping group? White-winged choughs have found a way by playing pied pipers and it is and remains the most unusual behaviour by any standard. The white-winged chough is possibly the only avian species worldwide that has such a mechanism for stealing juvenile youngsters from other groups. White-winged choughs are relatively easy to raise as nestlings but difficult to reintegrate into the wild. In a research project over 4 years in the 1980s, 15 cases were documented (Heinsohn 1991a) and my own observations in a different region stem from 2003, hence the behaviour may be relatively widespread. The abduction is sometimes based on in-group fighting but can also be achieved by a dazzling display (Fig. 10.4) that implies the existence of plentiful food. Essentially, the adult performs the elaborate display and the mesmerised youngster is readily led away. The interesting part is that the youngsters then grow up and stay with the adopted family and, in these cases, so far not considered in general theoretical frameworks of cooperative behaviour, the youngsters are entirely unrelated but will eventually also grow into taking roles within the group.
The butcherbird has even more sinister thoughts when playing pied piper. The observation was made outside Armidale, New South Wales, near an outdoor aviary containing 13 budgerigars. Their flight aviary was half covered with shade cloth and the entire aviary covered in chicken wire (small mesh). I heard the song of a pied butcherbird and found it sitting on top of the aviary section without shade cloth and singing. I noticed that one of the budgerigars flew towards the butcherbird and hung on the wire very near the singing bird. Within a period of 15 minutes at the most, all 13 had assembled within a radius of a maximum of 300 mm away from the butcherbird. They were motionless as if mesmerised. The butcherbird hopped a little closer to the cluster of birds but, by that time, I sensed its intentions and went over to the aviary. The butcherbird only flew away once I entered the aviary. I discovered that a small hole had been cut into the chicken wire on the top. I concluded that the butcherbird was singing to entice them, or at least one of them, to get near that opening and thus get a rich bounty of a well-fed bird for a meal. This was the first and only time that I had observed that song can be used to lure another species into a trap. The behaviour of the budgerigars was remarkable. They had interrupted their usual range of activities. If they had been frightened they would have been able to take cover in many areas of the aviary. Instead, they all moved as close as possible to the singer and hung on the wire, belly up, exposing themselves to mortal danger. Clearly, the budgerigars were entirely unaware of the trap that was being set for them.

Fig. 10.4. Abduction dance. (Top) An adult white-winged chough fans its tail and stretches out its wings while slowly walking in a half circle, the fledgling watching. (Centre) In the next phase the tail is raised and the outstretched wings are pointed forwards so that the leading edge is almost touching the ground. (Bottom) In this very contorted posture, the adult turns back to the juvenile for a full vertical display of both wings as well as the raised tail now facing the juvenile. Wing displays in white-winged choughs can mean that there is plenty of food. Displaying in this fashion makes the juvenile follow the adult and the abduction is complete (taken from individual video-frames by author at some distance away from the scene, hence the poor quality but worth documenting).
Awareness of intentions of others
Evaluating behaviour in others may involve awareness. Such awareness of another’s state of mind can include future intentions and the budgerigars clearly had no idea what the butcherbird’s future intentions were. However, it has already been shown that it is not impossible for some birds to have a sense of future intentions in another (for instance, the blue jay that rehides its cache because it had seen that it was being watched). Again, we have no studies of Australian birds that provide robust evidence of a theory of mind in any species. However, there are occasionally encounters with birds or observations of birds that suggest awareness of another’s intentions and at least a considered assessment of a situation.
There are sometimes glimpses to be had about the way birds deal with new and unfamiliar situations. For instance, a little eagle (Hieraaetus morphnoides) was once delivered to me in very emaciated condition. Examination revealed that it suffered from crop impaction and was unable to feed or drink. Crop impactions block the food pipe and the crop itself can get infected. The experience would also be very painful for the bird, not to mention that full compaction leads to a slow and painful death. The methods of treatment are limited, expensive and can carry substantial risks. One very simple method, not always successful, is to fill a syringe with lukewarm water mixed with liquid paraffin, affix it to a short plastic tube, then feed the tube slowly down, and then slowly introduce the fluid into the food pipe. This is done to soften the impaction but it needs continuous, twice-daily treatment and has to be performed on a conscious bird so it can participate with some swallowing if possible. The eagle had been delivered directly from the wild and a few hours later, I was to give it its first treatment. I wear a welder’s leather apron for such occasions so the bird can comfortably sit on my lap reducing risk of injury from the talons.
Before I began the procedure, I showed the eagle the ‘instruments’, then opened its beak while someone else was ready to hold the wings in case the bird began to flutter but it was otherwise unrestrained, and I gently inserted the tube and delivered the fluid. By nightfall, the procedure had to be repeated. I again sat the bird on my lap, then showed the instruments to the bird.
To my very great surprise the eagle did the following: it sat quietly, made no attempt to move or escape, then it slowly turned its head, first looking straight at me (with the beak about 5 inches away from my face), then looked at the instruments displayed on the couch and ready to use – for which action it extended its neck so that the ‘looking’ seemed very deliberate. What followed left me speechless: the eagle then turned its head back to a position parallel to mine, lifted its head, tilted it back a little and then opened its beak! Without any restraint whatsoever, the fluid was administered in the same manner and in the following days I was able to repeat the procedure eight more times with the eagle displaying the same behaviour. To my delight, the treatment was beginning to work and, once the impaction had cleared completely and the bird was able to feed normally and regain some weight, it was released. The behaviour that took me by surprise was the voluntary participation of the bird in its own treatment and the swiftness with which the bird had comprehended the point of the procedure. Obviously, one can say that the lack of objection stems from the first experience, which might well have provided some pain relief. However, the entire series is not explicable merely in terms of pain relief. It required a recognition of the instrument and some understanding of the sequence of events and, dare one say, intentions. Opening the beak while tilting the head backwards was an untrained intentional movement and the fact that it was repeated on a number of occasions, and only in this specific treatment context, makes this a deliberate, not incidental, action.
Equally, I had to treat a wedge-tailed eagle, having to administer daily injections twice a day into the leg of a fully-grown adult male, and needed to do so on the conscious bird. I used a sawing horse, wrapped the eagle into a large woollen blanket (to stop the powerful wings from flapping ), provided a pillow for its head and then laid down the bird on its back and from that position it could comfortably watch the procedure. My assistant held the other leg, but this was just a precaution against the powerful talons. Every injection proceeded without the slightest incident but the eagle was allowed to watch and did so without fail throughout every procedure and without fighting it. Again, this was a wild eagle. What can one assume from this behaviour in both these eagle encounters? Was insight involved in these instances and did the birds understand that they were being helped? It was clear that in each case I tried to indicate what I was about to do and did so slowly and deliberately and gave the bird a chance to follow the sequence of events. It is difficult to fathom the processes in the bird’s head. Both procedures were invasive and the injections would have caused minimal pain, but pain nevertheless.
It would appear that there might be many opportunities and rich offerings to test cognition generally in Australian birds further and also examine the merits of the social brain hypothesis in pair-bonded birds and in cooperatively breeding species. As their social life can be very complex and may involve siblings and unrelated outsiders their need for communication is likely to have become more demanding. In other words, in the field of communication among Australian birds, one can find a wide range of solutions to the question of regulatory mechanisms for pair and group living, ranging from clever adaptations to very complex behaviour requiring advanced cognitive abilities, the latter probably fostered by the need for cooperation in the first place.