Chapter Nine
INTRODUCTION
The protozoa is a diverse assemblage of single-celled organisms found in most aquatic environments sustaining life. Foraminifera are shell-bearing (i.e., test) rhizopods (Order Foraminiferida), that are one of the most abundant groups of small marine and estuarine protozoa (Phleger 1960; Lee 1974; Be 1977; Boersma 1978). In addition to the foraminifera, two other important groups of marine protozoa are the siliceous radiolarians (Superclass Actinopodea) and the ciliated tintinnids (Order Choreotrichida). While the foraminifera are not usually considered among the major "constructors" of Holocene reefs (Fagerstrom 1987), they are nonetheless one of the most abundant coral reef organisms, with a capacity to produce enormous quantities of biogenic skeletal material, which may eventually be consolidated into the reef framework (Muller 1974; Sakai and Nishihara 1981; Salvant and Venec-Peure 1981). Benthic foraminifera may play an important "reef-building" role on a number of large (>10 km2) oceanic platform reefs (e.g., Lucipara Islands, Karang Skaro, Nil Desperandun, Karang Dusborgh, all in the Banda Sea), where they seem to contribute a significant amount of biogenic carbonate to the construction of extensive reef flats. However, there are no quantitative studies available to support this recent field observation. It is interesting to point out, however, that, unlike many other oceanic reef systems (e.g., Palau, Hawaii, etc.), the Banda Sea coral reef platforms are located in an area of intense seasonal upwelling (Wyrtki 1961), with correspondingly high productivity (Gieskes et al. 1990)', and more importantly, an abundant supply of inorganic nutrients which are pumped into the euphotic zone during each Southeast Monsoon (Wetsteyn et al. 1990). Along the coastlines of large islands fringed by reefs (e.g., Bali), the skeletal remains of benthic foraminifera are a significant source of beach sand, thus of considerable economic value to regional economies (fig. 9.1).
Hallock (1976) documented earlier that approximately 0.2% of the coastal and nearshore sand reservoir of Oahu, Hawaii, was produced by the benthic symbiont-bearing foraminiferans Amphistegina, Heterostegina and Marginopora, with Amphistegina accounting for about 90% of the total production. Contribution of benthic coral-reef foraminifera to the total primary production of the reef system has not been fully determined, but their role as primary producers must be significant. In Palau, carbonate production estimates of rotaliinids on the high-energy seaward reef flats were up to 2.8 kg CaCO3.m-2.yr1, while on the lagoonal slopes the productivity stood at 0.6 kg CaCO3.m-2.yr-1 (Hallock 1981b). These, and other early studies, clearly demonstrated the significant role of reef-associated benthic foraminifera in the production of reefal sediments (Chapman 1900). However, their contribution to reefal build-ups has been generally unappreciated, even though the group had a significant input in reefal development since the late Paleozoic (Hallock 1981b). Extensive limestone deposits on Java, Madura, Sumatra, and the impressive Neogene limestones of Mangkalihat Peninsula in East Kalimantan, for example, are of benthic foraminiferal origins. In Bali, the magnificent white cliffs along the azure coast of southwest Bukit Badung, near the ancient Ulu Watu Temple, are made mostly from early Neogene foraminiferal and coral limestone deposits, and together with the magic of Ulu Watu Temple, offer tourists (and the intrepid scientist) an unforgettable experience (fig. 9.2).
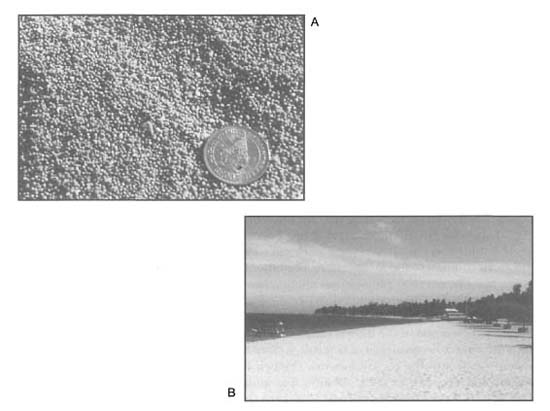
Figure 9.1. A) The beach deposits along the Sanur and Nusa Dua coastlines, southern Bali, consist almost entirely of the remains of benthic foraminifera, Baculogypsina sphaerulata. B) B. sphaerulata is a spherical benthic foram that thrives at the seaward edge of the Sanur reef flat.
Photos by Tomas and Anmarie Tomascik.
With regards to Recent reefs in Indonesia, it seems that corals, calcareous algae and benthic foraminifera are the major carbonate producers, a fact clearly recognized by Molengraaff (1928) and Wells (1957) in other coral reef systems, past and present. The role of benthic foraminifera in the construction of the coral reef framework was demonstrated by Finckh (1904) on Funafuti Atoll cores, in which, in terms of reef-building importance, benthic foraminifera were outranked only by the coralline algae and Halimeda spp. Surprisingly, scleractinian corals were ranked fourth. Scoffin and Tudhope (1985) have since shown that on the outer reefal shelf of the Great Barrier Reef, Australia, benthic foraminifera are the dominant producers of calcareous reefal sediments. Similar conditions seem to exist on most offshore and oceanic reefs in the Indonesian Archipelago.

Figure 9.2. Massive, early Neogene, limestone cliffs along the Bukit Badung (Bali) southern coastline are made almost entirely from foraminiferal and coral deposits raised by tectonic uplift.
Photo by Tomas and Anmarie Tomascik.
A vast majority of extant foraminifera are benthic; however, some families are primarily pelagic. There are less than 100 extant pelagic foraminiferan species (Murray 1991b). Nevertheless, they are very abundant and constitute a significant biomass of the zooplankton community, as well as being the most important contributors of CaCO3 to deep-sea sediments. About 30 planktonic foraminifera species are restricted mainly to the subtropical and tropical oceans (e.g., species of the Globorotalia, Globigerinella, Globigerinoides, Pulleniatina, Hastigerina); however, very little information is available on their biology, ecology and distribution in Indonesian waters. Most of the tropical pelagic foraminifera found in the surface waters (0100 m) are symbiotic with photoendosymbionts (e.g., dinoflagellates, mainly Gymnodinium beii, and Chrysophyta) (Spero 1987; Faber et al. 1988, 1989), and are therefore generally restricted to the euphotic zone (maximum depth about 150 m). A similar symbiotic association and distribution are found in the shallow-water benthic foraminifera, such as Calcarinidae, Asterigeridae, Alveolinidae, Amphisteginidae, Nummulitidae, and Soritidae (Hallock 1984). Most planktonic foraminifera live in the euphotic zone, and attain their maximum densities in the deep chlorophyll maximum layer (Murray 1991b), whose depth varies along both spatial and temporal lines (Fairbanks and Wiebe 1980). Non-symbiotic foraminifera, pelagic and benthic, on the other hand, have a much wider depth distribution, since they do not require sunlight for growth and development.
CLASSIFICATION
In shallow tropical coastal-water environments where high temperatures predominate, and seawater is supersaturated with respect to CaCO3, skeletal remains of microscopic calcifying organisms accumulate in great abundance, and foraminifera tests abound. The foraminifera of the Indonesian Archipelago are a diverse group of pelagic and benthic (intertidal to abyssal) protozoans found in marine and estuarine environments. Even though the extant foraminifera are among the most abundant groups of organisms in many benthic communities, they have received relatively little attention in Indonesia, mainly because of taxonomic difficulty. The fossil record of foraminifera dates to the Cambrian, with over 35,000 species described. The exact number of extant species is not known, but is most likely somewhere between 3500-5000. Thus, foraminifera are one of the most widespread and long-lived groups of organisms in today's oceans.
To obtain a perspective of the taxonomic complexity of this group, consider the 70-plus volumes of the Catalog of Foraminifera. The first significant work on the extant marine benthic foraminifera of the Indonesian Archipelago was that of Hofker (1951), who described 16 families, 52 genera and 104 species from the Siboga Expedition collections. Twenty-seven years later, the benthic samples from the 1929-1930 Snellius Expedition yielded an additional 462 species (Hofker 1978). However, the vast majority of the Siboga and Snellius specimens are from oceanic deep-water samples, thus coral-reef-associated foraminifera are under-represented, mainly as a result of sampling effort. This diverse group of protozoans is "currently" assigned to the Kingdom Protista (or Protocista) (table 9.1). However, the taxonomic certainty of many of these groups is questionable at best, and the final fate of the Subkingdom Protozoa remains to be decided. As of now, the Protozoa is: "…a large, unwieldy assemblage, so grouped only by consensus of protozoologists" (Leeetal. 1985).
The foraminifera have characteristics such as cytoplasmic organization and pseudopodia that are very similar to most amoeboid organisms grouped in the Protozoa. One of the main characteristics that sets foraminifera apart from amoebae is the presence of hard biogenic tests, which most species secrete themselves, and from which the pseudopodia, or rhizopodia, protrude in a net-like array (Anderson and Lee 1991) (fig. 9.3). The extant foraminifera are currently grouped into four suborders and 13 superfamilies (table 9.2), but detailed classification of foraminifera is beyond the scope of this book, and the reader is referred to Ellis and Messina (1965, 1966) for an introduction to the foraminiferal systematics.
BIOLOGICAL ASPECTS
Test Morphology
The classification of Foraminiferida is based mainly on test morphology (i.e., architecture and ornamentation), growth patterns and material composition (Lee et al. 1985). Compared to other protozoa, foraminifera are relatively large organisms with adult test diameters ranging from < 0.1 mm (e.g., some species of planktonic foraminifera) to 100 mm (e.g., Marginopom vertebralis) (fig. 9.4). Forams may have life spans ranging from weeks to years (Lee et al. 1985; H. Spero, pers. comm. 1996). Foraminiferan tests are either simple unilocular (i.e., single chamber -Allogromiina) to relatively complex multilocular (i.e., many chambers - Rotaliidae) forms. In multichambered species, test growth is always initiated from a pro-loculus (i.e., the first chamber) to which new locula are added, each connected with the other by a foramina (thus the name foraminifera), which is the previous aperture (fig. 9.5). The locula (i.e., chambers) are added in a specific sequence depending on the species (e.g., rectilinear or coiled). The locula can be added in a variety of ways (uniserial, biserial, or triserial), and can produce a diversity of shapes from spiral, flat coiled, conical, etc. In many species the surface of the test is covered by elaborate projections or ornamentations, which are used for taxonomic identification.
Table 9.1. Current taxonomic position of Order Foraminiferida.
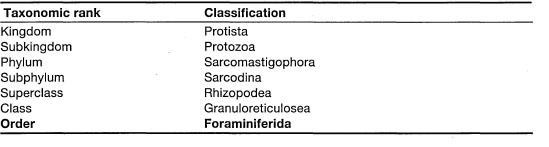
Table 9.2. Order Foraminiferida. List of suborders and superfamilies with some basic morphological characteristics.
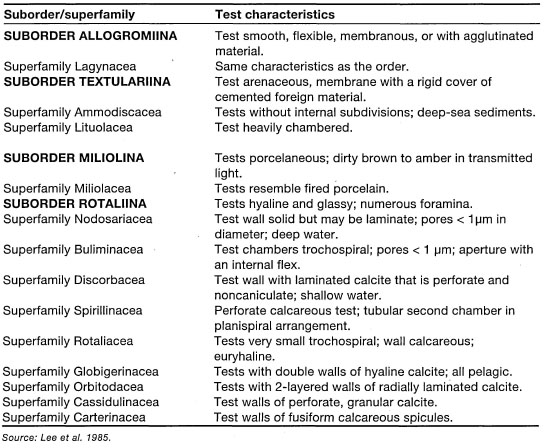
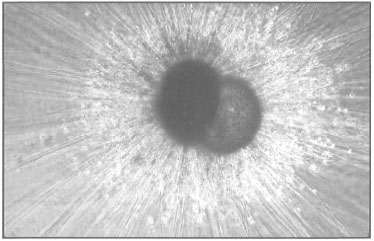
Figure 9.3. Photograph of a planktonic foraminifera Globigerinoides ruber, showing multi-chambered test with long delicate spines sheathed with cytoplasm.
Photo courtesy of H.J. Spero, University of California, Davis.
The tests consist mainly of CaCO3, predominantly calcite, which can be hyaline (i.e., transparent), porcelaneous, granular, fibrous or alveolar (Lee et al. 1985). The extant benthic foraminifera with characteristically mineralized calcareous tests are the Suborders Miliolina and Rotaliina (Lee et al. 1985). However, foram tests may also be gelatinous, chitinous, arenaceous (e.g., Textulariina) or siliceous. Superfamily Robertinacea are characterized by perforated tests made of aragonite (e.g., Mississippina, Robertina, Rubratella).
In terms of surface features, foram tests can be either perforated by numerous small pores (i.e., perforations), or be imperforate with a distinct smooth porcela neous appearance (e.g., Qinqueloculina, Triloculina). In species without perforations, the pseudopodia stream out through a large aperture. In perforated groups, the small pores in the test allow the cytoplasm to stream out, thus forming the characteristic reticulopodia which appear to be in constant motion. The fine, filamentous pseudopodia (i.e., rhizopodia) which branch freely and anastomose with one another, exhibit an interesting phenomenon (a continual puzzle for the biophysicists), which is the bi-directional transport or "streaming" of small granules in the cytoplasm. It seems that the granules along the outer surface of the pseudopodia are always moving towards the centre (i.e., inwards), while the particles in the inner core of the pseudopodia are usually moving towards the distal (i.e., outer) portion of the pseudopodium. The sticky pseudopodia are highly efficient food traps. In symbiotic species, the photosymbionts contained in the cytoplasm may undergo diurnal migrations along the length of the pseudopodia. During daytime the photosymbionts are concentrated towards the outer periphery of the pseudopodia, while at nighttime they withdraw into the test. In some foraminifera, the photosymbionts are periodically "harvested" (i.e., digested) by the animal.

Figure 9.4. Marginopora vertebralis is a significant producer of reefal CaCO3 on many Indonesian reefs. Pulau Sago, Banggai Islands (depth 10m).
Courtesy of Coral Cay Conservation Ltd.
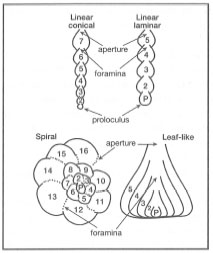
Figure 9.5. General patterns of test formation in Foraminiferida.
After Jahn et al. 1979.
REPRODUCTION
According to Lee et al. (1985), reproduction in foraminifera can be through gametogenesis, binary fission, budding, fragmentation or cytotomy (or plasmotomy where cytoplasmic division takes place independently of nuclear division in multinucleate protozoa). However, these modes of reproduction are restricted mainly to the smallest of the foraminifera (e.g., Allogromia spp.). Whatever the mode of reproduction, the initial growth of all foraminifera begins with the formation of the first chamber called the proloculus (fig. 9.6). In the unilocular species, the single chamber persists; however, in multilocular species new loculi are added in a species-specific sequence.
The majority of foraminifera have relatively complex life cycles involving dimorphic alternation of generations (i.e., asexual and sexual phases). Indeed, one of the most distinctive features of foraminifera is their mode of reproduction. Many foram groups exhibit dimorphism (or trimorphism), consisting of a megalospheric haploid sexual phase (i.e, gametic), and a microspheric diploid asexual phase (nongametic) (fig. 9.7). In fact, the life cycle of the foraminifera is highly complex, with eight different life-cycle programs recognized (Lee et al. 1991). However, of the 38 species studied in detail thus far, about 75% exhibit a metagenic life-cycle programme, which involves regular alternation of haploid sexual and diploid asexual generations (Lee et al. 1991). Only about 25% of foraminifera studied thus far exhibit an apogamic life-cycle programme, which is characterized by the predominance of asexual reproduction, and where sexual reproduction is either suppressed or does not occur (Lett et al. 1991). A gamic life-cycle programme, where sexual reproduction predominates, has been conclusively demonstrated in all planktonic forams studied so far (e.g., Orbulina universa, Globigerinoides ruber, G. sacculifer, G. conglobatus, Globigerina bulloides, Hastigerina pelagica) (H. Spero, pers. comm.).
The mode of sexual reproduction varies as well, with about 70% of studied species undergoing gametogamy, characterized by the fusion of haploid gametes from two or more gamonts with subsequent formation of zygotes (Lee et al. 1991). Only about 10% of sexual reproduction occurs through autogamy, which involves the fusion of two haploid nuclei from the same parental cell (Lee et al. 1991).
The uninucleate haploid gamonts represent the sexual phase of the reproductive cycle in foraminifera. The maturation of the gamont results in the formation of flagellated gametes which are released into the sea where fusion with gametes from other gamonts takes place. Thus zygotes are produced externally, and the release of gametes by the gamont is analogous to broadcast-spawning in corals and other reef invertebrates. Not much is actually known about the planktonic phase of the zygote; however, it is well-equipped to survive in the plankton. The maturing zygote forms a microspheric proloculus and the animal begins to undergo intracellular nuclear divisions, with successive additions of new chambers. The asexual phase of the life cycle is diploid. The multinucleate animal in the asexual phase is called an agamont. In holoplanktonic species agamont maturation is completed in depths of less than 100 m. Once the agamont matures meiosis occurs, thus resulting in the formation of mononuclear cells called agametes which upon maturation are released from the agamont. The maturing agamete forms a megalospheric proloculus around itself, and successive additions of new chambers and gamete maturation begin once again. This reproductive sequence is a highly simplified generalization of the life cycle of most foraminifera. According to Lee et al. (1991), only about 38 species have been studied in detail thus far. Note, however, that there has never been any direct observation of asexual reproduction in any of the planktonic foraminifera (H. Spero pers. comm.).
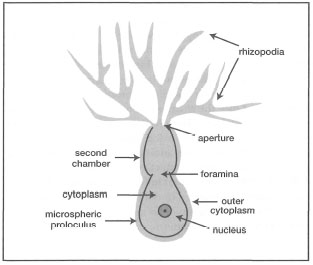
Figure 9.6. General morphology of a juvenile foraminifera.
Source: Jahn et al. 1979.
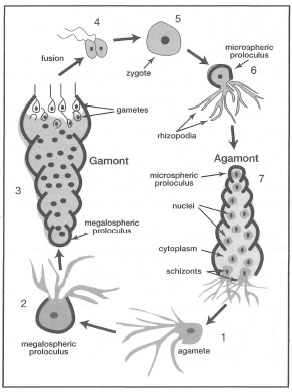
Figure 9.7. Generalized representation of the metagenic life-cycle programme with alternation of haploid sexual and diploid asexual generations in foraminifera. 1) uninuclear gamont or agamete; 2) juvenile gamont with megalospheric proloculus; 3) mature gamont with pregametogenic mitosis followed by pregametic nuclei; maturation of isogametes; 4) fusion of gametes from different gamonts which usually form "nuptial cysts"; 5) diploid zygote; 6) juvenile agamont with developed microspheric proloculus; 7) mature agamont with simultaneous meiotic divisions followed by cytokinesis and formation of gamonts by schizogony (gamogony).
Based on Beck and Braithwaite 1968.
SYMBIOSIS
Considering the extent and diversity of algal endosymbionts in foraminifera, our previous neglect of the phenomenon is difficult to put into perspective… Their abundance in today's seas and their contribution to CaCO3 shell production is not generally appreciated.—LEE AND ANDERSON 1991
The foraminifera are hosts to an amazing variety of photoendosymbionts that includes dinoflagellates, chlorophytes, unicellular rhodophytes, diatoms and chrysophytes (Lee et al. 1985; Lee and Anderson 1991). One of the most interesting recent discoveries was that certain groups of temperate foraminifera, notably the Elphiidae, Rotaliellidae and Nonionidae, are able to retain and use (i.e., husband or farm) chloroplasts from some algae which they have partially consumed (Lee and Anderson 1991); however, this has so far not been reported in tropical species. Most of the tropical symbiotic foraminifera are larger pelagic and benthic species. Size is relative, however, and in the world of Protozoa, foraminifera are the giants. Coral reefs in particular provide a variety of warm, shallow- and deep-water habitats which are favourable for the growth of photosynthetic benthic foraminifera (table 9.3).
The three dominant groups of benthic symbiotic coral reef-associated foraminiferans are the Families of Soritidae, Alveolinidae and Calcarinidae. This interesting and diverse group of coral reef organisms has, however, attracted relatively little interest from the biological community in Indonesia, even though knowledge of their life history strategies and environmental requirements would be of considerable benefit to petroleum geologists. Studies on benthic and pelagic foraminifera in Indonesia have only covered the thanatocenoses, and most research on this group has been done by geologists. It is interesting to point out that in the special reports published by the Lembaga Ilmu Pengetahuan Indonesia (LIPI), all foraminifera papers are assigned to the geology section.
The Family Soritidae are a common group of shallow-water benthic foraminifera abundant along many tropical coastlines as well as coral reefs islands and atolls. Of all the invertebrates, soritid foraminifera are hosts to the most diverse assemblage of photoendosymbionts. The photoendosymbionts belong to such diverse groups as dinoflagellates, unicellular chlorophytes and rhodophytes (Lee et al. 1985). Among the most important coral-reef-associated genera are Peneroplis, Marginopora and Sorites. Family Alveolinidae, with their distinct cigar-shaped tests are a common component of the coral reef community, with genus Borelis being host to diatom-like photoendosymbionts (Lee et al. 1985). Diatoms are characterized by the presence of siliceous frustules, which, however, do not form when the diatoms are endosymbiotic. It seems that the suppression of frustule formation while in the host's cytoplasm is associated with the endosymbiotic lifestyle, since when removed from the foraminifera the diatoms are able to form normal frustules again. Scientists have been successful in artificially culturing diatom-like endosymbiont isolates (Lee et al. 1979b) from a number of benthic reef-associated foraminifera (e.g., Heterostegina depressa, Amphistegina lessonii) (Lee et al. 1980b), which resulted in the formation of characteristic siliceous frustules, thus enabling identification of a number of new species of pennate diatoms (e.g., Fragilaria shiloi and Navicula reisii) (Lee 1980a). Since this early work considerable progress has been made and research has expanded into the Indo-Pacific. According to Lee et al. (1992), there are about 20 diatom endosymbiont species recognized thus far. In a study conducted in Palau, Lee et al. (1992) reported that the most abundant endosymbiont in the eight species of foraminifera studied was Nitzschia frustulum var. symbiotica, which was found in 24% of all samples collected. Other diatoms isolated from the Palau samples were Amphora erezii, A. roettgeri, Fragilaria shiloi, Cocconeis andersonii, Navicula sp., N hanseniana, Nitzschia laevis, N panduriformis var. continua, and Achnanthes maceneryae. Considering the close proximity of Palau to Indonesia, comparative study would be of interest. The eight species of benthic foraminifera from Palau that contain endosymbiotic diatoms are Amphistegina lessonii, A. lobifera, Heterostegina depressa, Neorotalia calcar, Calcarina spengleri, C. defrancei, C. gaudichaudi, and Baculogypsina sphaerulata. The Palau study conclusively demonstrated that foraminiferan hosts may contain a number of the endosymbionts at the same time. For example, Calcarina gaudichaudi houses nine endosymbiont species. In general it seems that most foraminifera contain between five to eight endosymbiont species at one time (Lee et al. 1992). However, Amphistegina lessonii and Heterostegina depressa each contain only three endosymbiont species. In Amphistegina lessonii the dominant endosymbionts are Fragilaria shiloi and Nitzschia frustulum ver. symbiotica, while in Heterostegina depressa the two dominant endosymbionts are Cocconeis andersonii and Amphora roettgeri (Lee et al. 1992). There also appears to be a geographic variation in the endosymbiont complement in the host animal. For example, Heterostegina depressa from the western Indo-Pacific (i.e., Palau and Kudaka Island, Japan) does not contain Nitzschia frustulum var. symbiotica (sample N=32). As was pointed out by Lee et al. (1992), inclusion of earlier studies revealed that both Amphistegina lessonii and Heterostegina depressa contain more endosymbionts as the sampling area increases. When considering the entire collection, H depressa is host to 12 endosymbiont species (Lee et al. 1992), thus there is a significant geographic variability.
Table 9.3. General distribution of known algal symbiont-bearing foraminifera on coral reefs and their preferred habitat (based on abundance). Note that the species listed may be found in more than one habitat.

The dominant group of benthic foraminifera in the Indo-Pacific are the rotaliines, especially Family Calcarinidae. Some of the most important endosymbiotic rotaliines are Amphistegina, Baculogypsina, Calcarina, and Heterostegina. Endosymbiotic diatoms are associated with both Amphistegina and Heterostegina. Families Cymbaloporidae (e.g., Cymbaloporetta) and Homotrematidae (e.g., Homotrema) are important coral reef-dwellers worldwide (Lee et al. 1985). In an excellent review of foraminiferan symbiosis, which the reader is urged to refer to, Lee and Anderson (1991) provide a list of all known foraminiferan-endosymbiont associations. In total, there have been 47 foram-endosymbiont groups studied, thus far (i.e., up to 1991). Of all the known symbiotic foram-endosymbiont associations about 38% involve diatoms (e.g., Nitzschia), 26% chrysophytes (not yet identified), 21% dinoflagellates (e.g., Gymnodinium), 9% unicellular chlorophytes (e.g., Chlamydomonas), and 6% unicellular rhodophytes (e.g., Porphyridium). What seems to be interesting from the coral reef perspective, is that the three main coral reef-associated groups, Alveolinidae, Amphisteginidae and Calcarinidae, harbour only Bacillariophyta (diatoms), while the abundant coral reef soritids (e.g., Marginopora) are symbiotic with dinoflagellates.
The evolution of symbiosis with a variety of photoendosymbionts has resulted in some significant morphological and physiological adaptations in the host species. Lee and Anderson (1991), and others, have asserted that: "algal endosymbiosis was, in fact, a driving force in their [foraminifera] evolution". One of the most interesting adaptations is the suppression of sexual reproduction. According to Bermudes and Back (1991), the suppression of sexual reproduction maybe related to the fact that asexual reproduction guarantees direct transfer of symbionts to the next generation, while sexual reproduction requires that the zygote, or the maturing agamont, must be reinoculated with the endosymbionts from the ambient seawater. Whether some gametes contain endosymbionts when they are released from the gamont is not known, and is an interesting field of research. It appears that in the case of foraminiferan symbiosis, each biont (i.e., partner) may live independently, thus the symbiosis is facultative (Bermudes and Back 1991). The suggestion that the relationship may be ecologically obligate (Bermudes and Back 1991) may be too wide a generalization, since there are extensive shallow-water tropical coastal areas where inorganic nutrient and food limitation is not a factor (e.g., near major rivers or in upwelling areas).
In general, symbiotic foraminifera tend to be much larger than their heterotrophic cousins, as is true for other symbiotic organisms, such as corals (Hallock 1985; Lee et al. 1979a; Hallock et al. 1991). For example, tests of Marginopora vertebralis can be over 100 mm in diameter. Other species such as the spherical Baculogypsina can reach over 2 mm during reproduction. With increased test size (i.e., body size) the sexual maturation has been delayed, compared to the purely heterotrophic groups, and according to past studies, their life spans are correspondingly longer (i.e., a few months to a year). However, some large deep-water foraminifera do not contain endosymbionts, while some very small foraminifera do (Hallock et al. 1991). According to Hallock et al. (1991), photoendosymbiosis and growth to a large size in foraminifera are considered adaptations to environmental conditions where food and energy resources are consistently limited. In comparison to the heterotrophic species, tests of benthic symbiotic foraminifera are significantly thinner and flatter, an adaptation that allows greater transmission of light, and exposes greater surface area to the downwelling solar radiation.
In contrast to the flattened test morphologies of benthic symbiotic foraminifera, planktonic symbiotic foraminifera have evolved thin, spherical tests with numerous highly perforated chambers (e.g., Globigerina bulloides, G. falconensis, Globigerinella calida, Globigerinoides sacculifer, G. fistulosus, Globorotalia menardii), and elaborate ornamentation (e.g., spines) along which cytoplasm may stream out. In many planktonic species the endosymbionts are "farmed", and move along the pseudopodia in response to daily light cycles (Be et al. 1977). In all symbiotic foraminifera the endosymbionts are housed in specialized vacuoles within the cytoplasm (i.e., they are intracellular) (Bermudes and Back 1991). According to Lee (1983), the symbionts can constitute a significant percentage of the holobiont biomass.
The above morphological adaptations seem to be directed at maximizing the photosynthetic efficiency of the photoendosymbionts as well as increasing the capacity for absorption of dissolved nutrients from ambient seawater. What nutritional role do the endosymbionts play in the association? Lee and Anderson (1991) point out that endosymbiosis in the larger foraminifera, with its tight recycling of nutrients and organic compounds, was an adaptation to oligotrophic tropical and subtropical oceans where they tend to predominate. It seems that the foram-endosymbiont system is functionally similar to the coral-zooxanthellae symbiosis; however, much less research has been done in this field (Lee et al. 1979; Hallock 1981a; McEnery and Lee 1981). Nonetheless, it seem clear that the foraminifera may potentially obtain a significant amount of energy from the endosymbionts in the form of various photosynthates that are translocated to the host animal, as is the case with the coral-zooxanthellae symbiosis. The potential amount of energy obtained by the animal biont is also dependent on the nature of the photoendosymbiont. For example, Hallock et al. (1991) pointed out that diatom-foram holobionts have higher rates of CO2 fixation (i.e., productivity) than do rhodophyte-foraminifera holobionts. What little information exists, suggests that in some associations (e.g., diatom-foram) the main photosynthates translocated from the endosymbiont to the animal biont are glucose and glycerol, which can be utilized by the animal biont in supplementing metabolic energy costs and/or growth (Keremer et al. 1980). However, some foraminifera, such as Heterostegina depressa, are apparently able to absorb sufficient nutrients from seawater so that they do not have to actively feed.
Calcification
In addition to the potential energy and food subsidy offered by the endosymbionts, the endosymbionts most likely have a considerable influence on the calcification process. Recent studies indicate that this may vary considerably among different species. For example, Ter Kuile and Erez (1987) demonstrated, using radionuclide C pulse-chase experiments, that Amphistegina lobifera was able to incorporate significant amounts of photosynthetically derived G into its skeleton, while a similar species was not. There have been a number of theories suggested in the past to explain the calcification process in animal-algal symbionts, and one of the earliest is that light stimulates calcification through CO2 fixation (i.e., photosynthesis). In this theory, the driving mechanism behind light-enhanced calcification in photoendosymbiont-bearing foraminifera is an increase in pH as a result of photosynthetic uptake of CO2, which increases carbonate concentrations in the animal tissue near sites of carbonate deposition. However, Ter Kuile (1991) points out that new evidence indicates that calcification in some algal symbiont-bearing foraminifera is not stimulated by the photosynthetic fixation of CO2. Nonetheless, a number of studies have shown that calcification rates of endosymbiotic foraminifera incubated in light were up to three times higher than those incubated in the dark (Lee and Anderson 1991; Lea et al. 1995). Another mechanism evoked is the organic matrix theory which suggests that an organic matrix initiates or inhibits calcification by spatially arranging the calcium and carbonate ions (Ter Kuile 1991). Recent experiments with Orbulina universa suggest that Ca2+ in this species is in direct isotopic equilibrium with surrounding seawater, and that less than 0.5% of shell Ca2+ can be attributed to another source (Lea et al. 1995). According to these authors, "an internal cytoplasmic Ca2+ pool probably does not provide a means by which seawater cation ratios can be fractionated". Furthermore, their experiments demonstrated that calcification rates in adult O. universa are 2-3 times higher under high light intensities than in the dark. Since the enhancement in calcification rate is close to the calculated increase in carbon ion due to the photosynthetic activity of the endosymbionts, Lea et al. (1995) pointed out that calcification rate in O. universa may be proportional to the degree of carbonate saturation.
Other possible mechanisms may involve active concentration of the reactants, since a number of foraminifera have been shown to concentrate calcium in internal pools (Ter Kuile 1991). One of the most plausible benefits of the symbiotic association for the foraminifers is the removal of metabolic wastes by the endosymbionts during photosynthesis. The removal of excess ammonium, phosphate and magnesium from the site of calcification will enhance the calcification process, since these substances are well-known crystal poisons. Whatever the final mechanism, the field has attracted considerable interest worldwide; however, up to now no comparable research has been initiated in Indonesia.
HETEROTROPHIC-FEEDING
Most pelagic and benthic foraminifera are heterotrophic organisms feeding on a variety of organic matter (i.e., carnivorous, herbivorous, detrivorous or omnivorous), which is available in the water column, or in the sediments. Planktonic foraminifera are generally omnivorous (Anderson et al. 1979; Be 1982), feeding on any organic material that may get ensnared in their sticky pseudopodial nets (fig. 9.8). They can be considered as passive suspension-feeders, since they rely on collisions with food particles such as zooplankton, phytoplankton or suspended organic particulate matter (Murray 1991b). Feeding is essential for the growth of planktonic foraminifera; however, they may survive without food for long periods provided photosynthesis of their endosymbionts is not inhibited (Caron et al. 1981).
Pelagic foraminifera are particularly well adapted for capturing zooplankton. The pseudopodial net of Globigerinoides sacculifer is highly effective in ensnaring small calanoid copepods (Spindler et al. 1984), which are its primary food source. The surface of their thin, perforated and multilocular tests is usually ornamented with long, thin and very delicate spines (often damaged in specimens from plankton tows), along which sticky cytoplasm can easily be seen streaming. In many species (e.g., Globigerinoides sacculifer, Orbulina universa) the pseudopodial nets supported by the spines substantially increase the surface area of the feeding surface. The spines can be very long, in as much as it is actually possible for scientists to drift in the water, and capture individual foraminifera (e.g., Globigerinoides spp.) in small glass jars as they drift by (Tomascik and Mah, pers. obs.). Capturing planktonic foraminifera in jars is a morning occupation of many foram scientists involved in experimental research, since great care must be taken so that the long delicate spines are not damaged, which would otherwise ruin the experiments.
Spinous planktonic foraminifera are especially abundant in oligotrophic water masses, where calanoid copepods are the dominant zooplankton. According to Spindler et al. (1984), planktonic foraminifera on an average capture and consume one zooplankter per day (fig. 9.9). During the Northwest Monsoon (i.e., non-upwelling period) the most abundant planktonic foraminifera in the central Banda Sea are Globigerinoides ruber and G. sacculifer, which are characteristic of oligotrophic water masses in other oceanic regions (e.g., Red Sea, central Indian Ocean) (Troelstra and Kroon 1989). During the same period the zooplankton community is dominated by copepods, which represent about 77% of zooplankton abundance (Baars et al. 1990). The oligotrophic conditions during the Northwest Monsoon are reflected by lower zooplankton biomass (9.5±2.8 cc.m-2) compared to the Southeast Monsoon upwelling season when the average zooplankton biomass in the eastern Banda Sea doubles (20.0±8.4 cc.m-2) (Schalk 1987). In more eutrophic regions, the planktonic foraminifera are dominated by non-spinose species (e.g., Neogloboquadrina dutertrei, Globorotalia menardii, Pulleniatina obliquiloculata), which are also mostly omnivorous. However, some species such as N. dutertrei feed exclusively on phyto plankton (Hemleben et al. 1980), and are usually associated with the Deep Chlorophyll Maximum layer where phytoplankton occurs in the greatest concentrations (Barmawidjaja 1993).

Figure 9.8. Photograph of a symbiotic planktonic foraminifera, Globigerinoides ruber, with ensnared zoo-plankton within the net-like pseudopodia.
Photo courtesy of H.J. Spero, University of California, Davis.
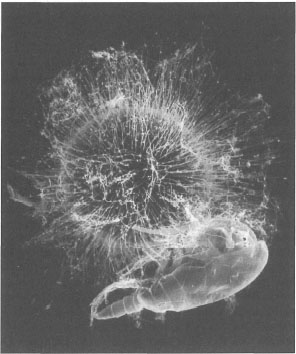
Figure 9.9. Photograph of a symbiotic planktonic foraminifera, Orbulina universa, feeding on a captured calanoid copepod.
Photo courtesy of H.J. Spero, University of California, Davis.
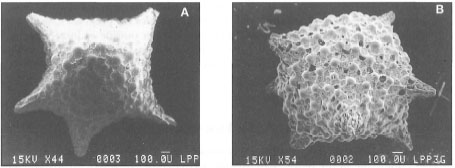
Figure 9.10. Benthic foraminifera are among the most abundant reef organisms. A) Micrograph of Baculogypsina sphaerulata (Parker and Jones), a dominant reef-associated benthic foraminifera at Sanur, Bali. B) Schlumbergerella floresiana (Schlumberger) at Nusa Dua, Bali.
Photos courtesy of M. K. Adisaputra, Pusat Pengembangan Geologi Kelautan -LIPI, Bandung.
Benthic foraminifera exhibit a variety of feeding strategies. Detritus-feeders (i.e., detritus plus associated bacteria) are present at all depths. Feeding occurs at the sediment/water interface or infaunally. Test morphologies in benthic foraminifera reflect feeding strategies. More erect types may be more common in environments where pseudopodial nets extended into the water column provide an efficient mechanism to trap suspended organic matter "raining" down, or drifting by in slow currents (Murray 1991a). Active herbivory in benthic habitats is restricted to the euphotic zone, where benthic foraminifera are known to forage (Travis and Bowser 1991). For example, Allogromia laticollarisi uses an extensive network of sticky reticulopods (i.e., highly branched and anastomosed filopods) that are used for locomotion as well as in accumulating benthic diatoms in net-like enclosures (Travis and Boswer 1991). Uptake of dissolved organic material may also be an important feeding strategy (Schwab and Hofer 1979). Many of the shallow-water foraminifera are symbiotic, thus translocation of photosynthates may be a significant supplement to their heterotrophic diets (Lee and Bock 1976). Many symbiotic benthic foraminifera in shallow-water coral reef environments are discoid or compressed, presumably to maximize photosynthetic efficiency of the holobiont. However, many highly abundant shallow-water symbiotic species, such as Baculogypsina sphaerulata or Schlumbergerella floresiana, are spherical, with a prominent spinous architecture, which provides an efficient mechanism for attachment to benthic algae in high-energy environments (seaward reef flat) (fig. 9.10).
Whether they are pelagic or benthic, feeding in most foraminifera is accomplished by the extrusion of pseudopodia (i.e., rhyzopodial nets) from the test of the animal. The spines serve as an efficient net in which zooplankton invariably are ensnared, and subsequently digested. The food particles (or prey) are engulfed by the cytoplasm and digestion usually takes place either outside or inside the tests with the assistance of digestive lysosomes. The particle size captured by foraminifera, benthic and planktonic, ranges between 1- 20 μm in diameter.
BENTHIC FORAMINIFERA
If there is one region of the planet that should have a high diversity of foraminifera, it is undoubtedly the Indonesian Archipelago. No other region in the world offers such a diversity of pelagic and benthic habitats as the thousands of islands and the open and enclosed seas of the archipelago. The pelagic habitats range from the warm oligotrophic surface waters to the cooler epipelagic nutrient-rich depths and the cold mesopelagic zone. In addition, vast areas of the Banda and Flores Seas are influenced by upwelling which results in highly productive surface waters during much of the year. In terms of benthic habitats, the Indonesian Archipelago offers a great diversity of shallow neritic environments (e.g., estuaries, mangroves, seagrass beds, coral reefs, etc.) as well as shallow-water oceanic habitats associated with coral islands, barrier reefs and atolls. Within the relatively small area of the Banda Sea, benthic foraminifera could be sampled from the sunlight intertidal reef flats of Pulau Manuk, and the dark hadal zone of the Weber Deep.
General Distribution
Among the best-known groups of foraminifera are the deep benthic forms, mainly as a result of sampling efforts during the past major expeditions. There are no comparable data on reef-associated foraminifera from the archipelago; however, benthic foraminifera from Jakarta Bay, Ambon Bay, Batam, etc., have been studied (Hamidjojo et al. 1980; Ongkosongo et al. 1980; Siregar and Hadiwisastra 1980; Helfinalis et al. 1989; Hermanto and Suhartati 1989; Suhartati and Subardi 1990). The available data indicate that some reefal foraminifera are represented in the sediments collected from bays fringed by coral reefs.
Hofker (1978) was one of the first to compare the distribution of the benthic foraminifera collected during the Snellius Expedition. He pointed out the great dissimilarity in benthic foraminiferan fauna from the western and eastern regions of the archipelago (table 9.4).
According to the samples collected during the Snellius Expedition, Hofker (1978) found that Jakarta Bay (western region) had only five benthic foraminifera common with samples from Sorong, Iran Java. The question is whether these differences are real (i.e., of ecological significance), or a sampling artifact. Furthermore, it is not possible to ascertain whether the sampling was done in comparable environments.
Helfinalis and Rositasari (1988) investigated the foraminiferan fauna of Pulau Pari, an offshore shelf platform reef with an extensive shallow-water reef flat and two distinct lagoons, one of which has a depth greater than 15 m. According to the classification of Hopley (1982), the reef complex of P. Pari may be viewed as a mature lagoonal reef. The reef supports a diverse foraminiferan fauna, which is distinctly different from the benthic foraminiferal assemblage of Jakarta Bay (table 9.5). However, because of different sampling techniques as well as different sampling efforts, the data in table 9.5 are not statistically comparable. Nonetheless, the data suggest that the differences in foraminiferan diversity among the various regions of the archipelago are most likely related to differences in environmental conditions of stations sampled. For example, the sediments of both Segara Anakan and Jepara are dominated by euryhaline foraminiferans, since both areas are influenced by high fresh water runoff. Coastal waters in these areas are estuarine (i.e., low salinities) during much of the West Monsoon (i.e., rainy season). To determine whether there are biogeographical patterns in the distribution of benthic foraminiferans, comparative studies are required. Coral reef-associated foraminifera offer a good opportunity for such studies, yet none have been done thus far.
ECONOMIC VALUE OF FORAMINIFERA
Foraminiferans have proved to be indispensable tools for paleoclimatic, paleoecological and paleoceanographic reconstructions. Van Bemmelen (1949) ranked Foraminiferida as the most important stratigraphic tools of the Tertiary in the Indonesian Archipelago. The fossil remains of foraminifera (and other calcareous and siliceous plankton) are important indicators in biostratigraphy and dating of marine sediments, and have assisted petroleum geologists in their search for new oil deposits. Foraminifera, and the limestone sediments which they produce in conjunction with other calcareous planktonic and benthic organisms, are also of considerable economic value, and have been so, since early human cultures developed around the world. Among the best-known marine limestone deposits in the western world are the famous white cliffs (actually dominated by coccoliths) of Dover in England, which are rivaled by the anthropogenically orchestrated limestone deposits along the Nile, namely the Egyptian pyramids. The main source of building blocks for the pyramids were Eocene nummulitic limestones, consisting mainly of the foram genus Nummulites. In Indonesia, mining of limestone is of considerable economic value, and many of the shiny marble floors that are currently highly popular, are an intricate mosaic of foraminiferal fossils from the distant past. For example, the fine marbles from Gunung Panggul (south coast of East Java; Panggul Bay) originate from Miocene limestone deposits consisting mainly of Foraminifera (e.g., Aleveolinella bontangensis) and corals (van Bemmelen 1949).
BALI CASE STUDY
The role of extant foraminifera in the development of beach deposits is of significant economic value, a fact that has unfortunately not been recognized in the past, or the present (but see Adisaputra 1991). One of the greatest attractions of tropical countries with coral reefs are the long, white, palm-lined beaches, which in combination with the sun, surf and the azure- to-turquoise-coloured clear waters are an irresistible magnet for millions of tourists from the temperate regions. Indonesia is no exception, and Bali in particular has recently experienced a tourism boom. Putting aside the fascinating cultural experience that one obtains from visiting Bali, much of the future success of the tourist industry will depend on maintaining the island's environmental integrity, especially of the coastal marine environments. One of the most pressing management tasks will be the maintenance of the beaches. Sanur's golden beach, which is one of the most valued attractions on the island, is composed almost entirely of bioclastics (i.e., sediments made entirely of broken fragments of biogenic skeletal material). Indeed, about 80% to 90% of Sanur beach sand consists of foraminiferal tests. The foraminiferal tests of various sizes are dominated almost entirely by the calcarinids Baculogypsina sphaerulata and Schlum bergerella floresiana, which contain photoendosymbiotic diatoms. As a result, they prefer shallow-water, high-energy coral reef habitats.
Table 9.4. Regional comparison of shallow-water Foraminiferida between Jakarta Bay (southwest Java Sea) and Sorong (Halmahera Sea).

Table 9.5. General distribution of some benthic foraminifera in the Indonesian Archipelago. 1) Jakarta Bay 1929-1930. Habitat is coral and mud; 2) Sorong, west Irian Jaya 1929-1930. Habitat is mud and sand; 3) Jakarta Bay 1975-1979. Habitat is coral and mud; 4) Batam, northwest Java Sea, Teluk Tering. Habitat sand; 5) Ambon Bay, Ambon, Moluccas, 1984-1987; 6) Segara Anakan, south Central Java. Habitat sand, mud, sandy mud; 7) Jepara, north Central Java. Habitat sand, sandy mud; 8) Pulau Pari, north West Java. Habitats - 8A: Reef flat, 8B: Lagoon; 8C: Reef slope.




SOURCE OF SANUR BEACH SAND
Where does the immense amount of biogenic carbonate come from? The answer to this question lies in an extensive coral reef complex that runs along the entire length of the Sanur and Nusa Dua coastlines. Coral reefs that fringe large stretches of Bali's coastline are the classical fringing reefs. Along the south coast, especially Sanur beach and Bukit Badung Peninsula (e.g., Nusa Dua), the fringing reef extends a considerable distance offshore (e.g., up to 700 m at Cemara beach). Because of its relatively long and unbreached length (i.e., 8 km long), as well as a wide boat channel (sensu Guilcher 1988) (i.e., lagoon or moat), the Sanur fringing reef complex has often been incorrectly referred to as a barrier reef. The seaward edge of the fringing reef is above the mean high-water neap mark, and is delineated by white breakers. The depth of the boat channel varies; for example, between Bali Hyatt beach and Cemara beach the bottom of the boat channel is roughly at the mean low-water spring mark, whereas just off the Bali Beach Hotel to the north, the bottom is at about mean low-water neap mark. Note that the tidal range along the south coast of Bali is about 2.6 m. Because of a relatively restricted connection with the sea, water level inside the boat channel during low tides is above the offshore sea level, and thus most of the boat channel is always flooded.
Baculogypsina sphaerulata and Schlumbergerella floresiana are large spherical foraminifera that flourish at the seaward edge of the reef flat, and especially at the outer reef crest. These zones are classified as high-energy environments, receiving the full force of the Indian Ocean swell, a fact much appreciated by the world surfing community. At the seaward edge of the reef (i.e., algal rim), both species live attached to benthic macrophytes (e.g., Eucheuma spp., Galaxaura spp., Codium spp., Gracilaria spp., Acanthophora spp., Rhodymenia spp., Lurencia spp., Tricleocarpa spp., Hypnea spp., Dyctiosphaeria spp., and Sargassum spp.). The calcareous rhodophytes, particularly Lithophyllum moluccense, Jania spp. and Amphiroa spp., also provide suitable substrate for the attachment of these two benthic foraminiferans. In addition, both Sanur and Nusa Dua boat channels are important seagrass habitats (i.e., eight seagrass species recorded: Enhalus acoroides, Cymodocea serrulata, Thalassia hemprichii, Thalassodendron ciliatum, Halophila ovalis, H. minor, Halodulepinifolia, H. uninervis) whose calm 'lagoonal' waters provide an ideal environment for a variety of other benthic foraminiferans. Among the most abundant benthic foraminifera in the seagrass habitats are Amphistegina lessonii (Amphisteginidae), Calcarina calcar (Calcarinidae) and Heterostegina sp. (Nummulitidae), which are symbiotic with Nitzschia frustulum var. symbiotica (diatom) (Adisaputra 1991; Lee and Anderson 1991). Other smaller shallow-water benthic foraminifera found along the southern coastline of Bali are Elphidium advenum (Elphidiidae), Quinqueloculina pseudoreticulata (Miliolidae), Q. seminulina, Q. parked, Spiroloculina sp. (Nubeculariidae), Textularia sp. (Textulariidae) and Cellanthus craticulatus (Adisaputra 1991). Both Textularia sp. and Elphidium advenum are euryhaline species capable of withstanding very low salinities. Greater abundance of these two species in the sediments along the southern section of Sanur beach is most likely related to the brackish water coastal habitats that dominate this region of the coastline. Nonetheless, their contribution to beach sediments remains much lower than B. sphaerulata and S. floresiana.
The composition of benthic foraminifera on the seaward slope of Sanur, Benoa and Nusa Dua reefs, is much different from the lagoonal habitats. According to Adisaputra (1991), offshore areas are dominated by foraminifera belonging to Orders Rotaliina, Miliolina and Textulariina. At depths of less than about 12 m, Cribrononion hispidulus (Elphidiidae) seems to be the dominant benthic foram (Adisaputra 1991), especially along the Benoa coastline, which is characterized by low salinities. Further offshore, species numbers increase with depth, and genera such as Triloculina, Spiroloculina, Ammonia, Articulina, Anomalina, Clavulina, Eponides, Brizalina, Miliolinella, Martinotiella, etc., become more abundant (Adisaputra 1991). Pelagic foraminifera do not contribute much to the beach sediments; however, they are present in offshore sediment samples at depths of more than 16 m. The most abundant pelagics are Globigerina bulloides, Globigerinoides cyclostomus, G. ruber, G. sacculifer and Neogloboquadrina dutertrei. Note that G. bulloides is an indicator of upwelling conditions, which occur along the south coast of Bali.
Much has been said about the degradation of the Bali reefs; however, rapid surveys of the Sanur and Nusa Dua fringing reefs revealed that the reefs are still in relatively good condition, and that with appropriate management of coastal activities the reefs will be able to survive, if not regenerate. Note that the relatively good 'health' of these reefs is mainly due to strong currents that continually flush the coastal areas and bring in clean oceanic waters. However, there are signs that major changes have occurred. The seagrasses in the lagoon have increased, and while this may not be a problem at this time, continual discharge of nutrient-rich waste-water (e.g., sewage) will eventually have an impact, as it did in many other areas. However, of greater concern to the beach stability is the ill-advised building of structures for beach protection such as groins and cement walls. Both Sanur and Nusa Dua beaches rely on the longshore transport system to continually replenish the beaches with dead tests (i.e., shells) of benthic foraminifera, which originate at the seaward edge of the reef crest. Interference with the longshore transport system will ultimately result in increased beach erosion, and subsequent loss of economic value.
FORAMINIFERA AS ENVIRONMENTAL INDICATORS
To obtain a better appreciation of Foraminiferida, and their significance in oceanographical and paleontological studies, we shall touch upon topics that may seem to be out of place in a section dealing with coral reefs. However, since foraminifera have played the key role in the unraveling of paleoclimates and paleoceanographic conditions, it seems appropriate, even though a deviation from standard practice, to discuss the various techniques used to extract environmental information from deep-sea CaCO3 deposits, especially foraminiferal tests. Studies on Foraminiferida have increased dramatically since the early 1970s with the realization of their value as tools in exploration geology (Hinte 1978) and as indicators of paleoclimatic (Blow 1969; Molfino et al. 1982; Barmawidjaja et al. 1989a, b; van der Kaars 1991) and paleoceanographic conditions (Vincent and Berger 1981; Ganssen et al. 1989; Charles and Fairbanks 1992; Whitman and Berger 1992). However, up to now there has been very little research conducted on the biology or ecology of benthic and pelagic foraminifera in Indonesia (Adisaputra 1989; Kleijne et al. 1989; Troelstra and Kroon 1989).
Species Associations
The co-occurrence of particular species groups of planktonic and benthic foraminifera as well as the presence of other planktonic microfossil groups (e.g., calcareous nanoplankton and radiolaria) is often used as an effective tracer of various water masses (Adisaputra 1989; Troelstra and Kroon 1989), as well as of past and recent environmental conditions (Troelstra et al. 1989). Close association of various species groups of foraminifera with specific water masses has proved in the past to be a very useful tool for paleoceanographic reconstructions (Be and Tolderlund 1971; Hemleben and Spindler 1983). This is possible because many species have specific environmental and ecological requirements. Temperature and nutrient concentrations are the two dominant environmental factors that seem to regulate the presence or absence of certain pelagic species groups. With regards to the distribution of benthic foraminifera, the primary environmental factor involved is depth (van Marie 1989).
Troelstra and Kroon (1989) have demonstrated that during the Northwest Monsoon (i.e., period of downwelling), planktonic foraminifera (28 known species) in eastern Indonesia can be grouped into five distinct (statistically derived) "associations", each characteristic of a specific region (fig. 9.11).
Note that, unlike other studies which use bottom sediment samples to identify distribution patterns of pelagic foraminifera, Troelstra and Kroon (1989) used surface water samples. The species associations identified by Troelstra and Kroon (1989) seem to be reflecting subtle changes in environmental parameters (e.g., sea surface temperatures and nutrients) that occur between the western Flores Sea and the eastern regions of the Banda Sea and Timor Trough. Adisaputra (1989), using the sediment-analysis approach, demonstrated that planktonic foraminiferal communities in the Flores Sea differ considerably from those found in the Lombok and Savu Basins (table 9.6). While some of these differences can be attributed to the depth of the lysocline and the carbonate compensation depth (see following discussion), Adisaputra (1989) concluded that the distribution of Neogloboquadrina dutertrei, which is the dominant planktonic foraminiferan in the Flores Sea, is largely a function of hydrological conditions. Using faunal analysis (i.e., changes in species assemblages) on sediments from a deep-sea core taken in the Molucca Sea, Barmawidjaja et al. (1993) were able to demonstrate that the continual presence of Neogloboquadrina dutertrei (herbivorous foram) since the last glacial suggests that the Deep Chlorophyll Maximum (DCM) layer has been a permanent feature in the region for the past 27,000 years.
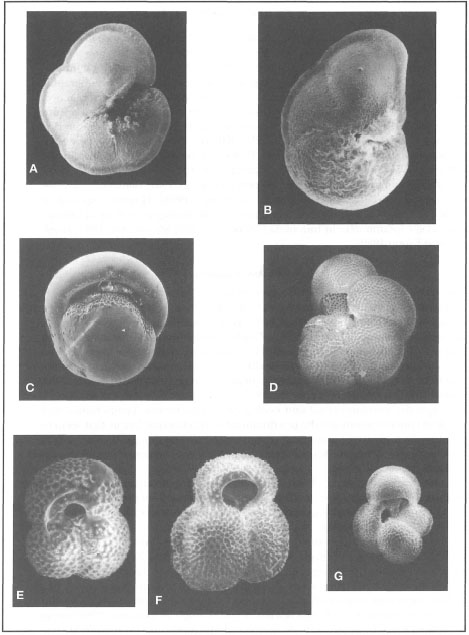
Figure 9.11. Recent planktonic foraminifera of eastern Indonesia from sediment cores taken in Flores, Savu and Lombok Basins. A) Globorotalia menardii; B) Globorotalia tumida; C) Pulleniatina obliquiloculata; D) Neogloboquadrina dutertrei; E) Globigerinoides cyclostomus; F) Globigerinoides ruber, G) Globigerina bulloides.
Photo courtesy of M. K. Adisaputra, Pusat Pengembangan Geologi Kelautan -LIPI, Bandung.
Of the 16 planktonic species recorded in the study, four species, namely Globorotalia menardii, Pulkniatina obliquiloculata, Neogloboquadrina dutertrei, and Globigerinoides ruber, were found in all stations above the lysocline. The planktonic foraminifera fauna in table 9.6 is characteristic of oceanic regions with well-developed DCM layers (Ravelo et al. 1990), and is very similar to the faunal composition of the north Molucca Sea deep-sediment core (Barmawidjaja et al. 1993).
In a parallel study, Troelstra and Kroon (1989) found that the Globigerinoides ruber, Globigerinoides sacculifer and Globorotalia menardii association (group composition 29%-58%, 12%-29%, and 7%-24%, respectively) seems to be characteristic of the open central Banda Sea surface waters, while the Globigerinoides ruber, Globigerinoides sacculifer and Neogloboquadrina dutertrei association (group composition 40%-60%, 8%-22% and 5%-16%, respectively) is characteristic of waters along the southern tip of Sulawesi and northeast Timor. The Banda Sea association, which is dominated by G. sacculifer and G. ruber, is characteristic of oligotrophic (i.e., low-nutrient) conditions known to exist in the region during the Northwest Monsoon, when surface downwelling predominates as a result of strong northwesterly winds (Wyrtki 1961). It is expected that surface distribution patterns of planktonic foraminifera will change significantly during the Southeast Monsoon, when upwelling generated by the strong southeasterlies predominates in the region. Troelstra et al. (1989) have in fact shown this to be true, by comparing surface abundance of planktonic foraminifera with the abundance of their tests from the surface layers of deep-sea sediment samples. The high abundance of Globigerina bulloides in the surficial layers of deep-sea sediments taken in the Seram and Tanimbar Troughs, two well-known regions of upwelling, and its virtual absence from the surface zooplankton community during the non-upwelling period (i.e., Northwest Monsoon), demonstrates conclusively that G. bulloidesis a good indicator of paleo-upwelling conditions (Troelstra et al. 1989). The high abundance of G. bulloides tests in deep-sea sediments indicates that productivity during the upwelling season must be considerable.
Subpolar Forams in Indonesia
Surprisingly, the subpolar, dextrally coiled, planktonic foraminifera Neogloboquadrina pachydermia has recently been recorded in the tropical waters of the Indonesian Archipelago (Barmawidjaja et al. 1993). This interesting discovery was made during the Snellius-II Expedition to the northern Molucca Sea. The main objective of the study was to obtain climatic proxy data for the reconstruction of climatic conditions in the northern Molucca Sea region during the last glacial period. Neogloboquadrina pachyderma was recovered from a deep-sea sediment core taken in the northeast sector of the Molucca Basin, about 50 km northwest of Halmahera. The sediment core was taken from a depth of 3510 m, and consisted of 533 cm of greenish clays with a rich and well-preserved foraminifera fauna as well as radiolarians and diatoms (Barmawidjaja et al. 1993). One of the most exciting discoveries was the great abundance of dextrally coiled N. pachyderma in sediment layers dated to the last glacial (i.e., 28-14 ka). The presence of N. pachyderma in the sediment core is rather unexpected, since it is a cold-water species. It was probably brought in by the intermediate depth current. Thus, its presence has some important paleoclimatic and paleoceanographic implications. According to Barmawidjaja et al. (1993), its occurrence in the northern Molucca Sea is most likely related to the nutrient-rich DCM layer, since it feeds exclusively on phytoplankton (i.e., a herbivore), as does its warm-water morphotype Neogloboquadrina dutertrei. N. dutertrei is a dominant planktonic foraminifera in the eastern Indonesian waters, as is indicated by the sedimentary record (Adisaputra 1989).
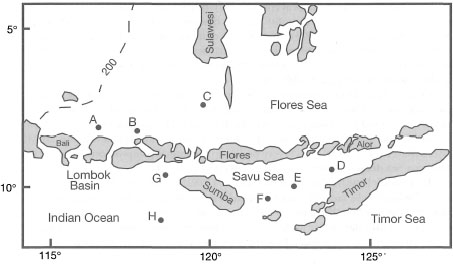
Figure 9.12. Map of Flores and Savu Seas showing locations reported in table 9.6.
Table 9.6. Abundance of some planktonic foraminifera tests in deep-sea sediment samples in Flores Sea: A (1537 m), B (1911 m), C (1500 m); Savu Sea: D (3295 m), E (3305 m), F (2514 m) and Lombok Basin (Indian Ocean) G (2085 m), H (4132 m) as for figure 9.12. Numbers in parenthesis are sample depths. Values in table represent percentages of the most common planktonic foraminifera in relation to total sediment composition. Data compiled from stations lying above the lysocline.
Deep Chlorophyll Maximum Layer
The Deep Chlorophyll Maximum layer is an ubiquitous feature of many tropical regions throughout the year, and is characterized by a significant increase in chlorophyll concentrations (i.e., up to a factor of 30). The high abundance of Neogloboquadrina dutertrei since the last glacial has been suggested as a proxy record for the presence of a well-established DCM layer in the eastern regions of the archipelago (Barmawidjaja et al. 1993). The depth of the present-day DCM layer in the northern Molucca Sea has not been established, but it is most likely associated with the nitracline (i.e., zone of maximum change in nitrate concentrations), and near the primary nitrite maximum (Parsons et al. 1984). In general, the DCM is located near the bottom of the euphotic zone, and, therefore, it should be located at depths of less than 100 m. Based on our reconstruction of vertical depth profiles (Oceanographical Cruise Reports No. 23 and 36) for temperature and nitrate during both upwelling and non-upwelling periods in the northeast Molucca Sea, it is suggested that the DCM layer is most likely located between the depths of 150 m to 75 m (fig. 9.13). Chlorophyll sampling of sufficient refinement to properly identify the DCM layer, which is only a few metres thick, is not currently available from this region.
However, Gieskes et al. (1989) were able to demonstrate that a well-defined DCM layer does indeed develop during the Northwest Monsoon in the eastern regions of the Banda Sea. The DCM layer becomes established in the thermocline during the non-upwelling period at a depth of 60 to 80 m (Gieskes et al. 1988). Figure 9.14 illustrates the development of a DCM layer in the east Banda Sea at Station 9 (04°39.2' S and 130°30.3' E) during February 1984 (Northwest Monsoon) and August 1985 (Southeast Monsoon) (Wetsteyn et al. 1990). Note, that the DCM layer has been observed to "spill" over offshore reefs as a result of strong tidally-induced currents or internal waves. The frequency of these events and their ecological significance has not been investigated.
The apparent upwelling event, indicated by the shallowing of the isopycnals (fig. 9.14A), is most likely associated with the intensification of the Mindanao Current (gyre) during the Southeast Monsoon. Strong southerly (i.e., from the south) monsoon winds that predominate in this region from July to October create strong northward-flowing surface currents from the northeast Molucca Sea with a net outflow into the Pacific Ocean (Wyrtki 1961). The net surface outflow from the northeast sector of the Molucca Sea is then balanced by upwelling of deeper nutrient-rich water along the west coast of Halmahera.
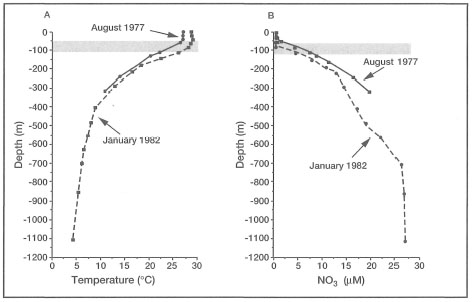
Figure 9.13. Temperature and nitrate vertical depth profiles for Station No. 11, Moluccas (Moluccas) Sea. Data compiled from Oceanographical Cruise Reports No. 23 and 36. Note the upward displacement of both the thermocline (A) and nitracline (B) during the upwelling period (Southeast Monsoon) in August 1977. The position of the Deep Chlorophyll Maximum (DCM) layer is suggested by the shaded areas.
Source: Institute of Marine Research 1977; National Institute of Oceanology 1982.
The analysis of the Molucca Sea sediment core revealed that at about 14 ka (at 270 cm depth), there was a pronounced shift in community composition of the planktonic foraminifera. The abundance of Neogloboquadrina pachyderma and Neogloboquadrina dutertrei (sinistral) increased, while there was a marked decrease in the abundance of the oligotrophic species group Globigerinella aequilateralis, Globigerinoides ruber and Globigerinoides sacculifer. Barmawidjaja et al. (1993) suggested that the marked increase of the two phytoplankton-feeding species with a concurrent reduction in the oligotrophic species group, indicates that the glacial DCM layer in the north Molucca Sea may have been considerably more productive than at present. However, the presence of the dextrally coiled Neogloboquadrina pachyderma is more problematic, and its origins remain highly speculative. New research efforts are required to solve some of these fascinating questions.
Morphological Indicators
The calcareous tests of planktonic and benthic foraminifera as well as a host of other calcifying marine organisms are excellent recorders of environmental conditions in which the animal or plant lived, and are, therefore, of particular interest to the paleontologists. One of the most pressing environmental concerns of the industrialized and non-industrialized world is the anthropogenically-induced global climate change, which according to many scientists is now inevitable (Houghton et al. 1990). The knowledge of past climates, paleoceanographic conditions and the reconstruction of ancient atmospheric CO2 concentrations are essential to our understanding of the environmental changes that may occur, and foraminifera have been in the forefront of this pursuit. These organisms have been used extensively in the retrieval of climatic data from the geologic record, which is essential for validation of current models in which oceanic temperatures are of paramount importance (Becketal. 1992).
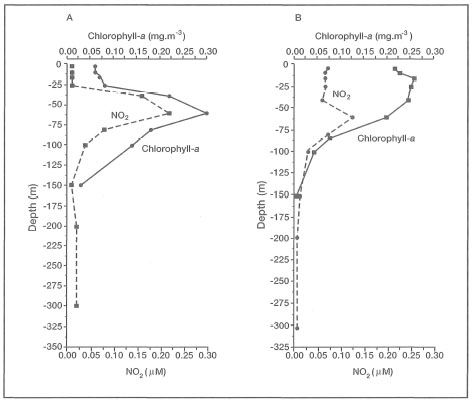
Figure 9.14. Deep Chlorophyll Maximum layer in the eastern Banda Sea (Station 9, 04°39.2' S and 130°30.3' E, Snellius-II Expedition) during: A) February 1985; and B) August 1984. Note that the DCM is associated with the primary nitrite maximum.
Source: Wetsteyn et al. 1990.
Test Porosity
Foraminifera have proved to be excellent indicators of environmental conditions, mainly because of their specific environmental and ecological requirements (Kennett 1976), and their relative ease of handling. Many groups of foraminifera exhibit well-defined intraspecific phenotypic differences (i.e., variations in test porosity, texture, thickness) which allow for separation of distinct sub-populations, each with subtly differing environmental requirements (Kennett 1976). One simple feature of foraminiferan test morphology that does not require elaborate analysis (only a simple microscope), and which has been of great assistance to paleontologists, is test porosity. Test porosity simply refers to the number of pores in the test; however, the value is presented as mean porosity which is the percentage of open pore area per unit area of test (Murray 1991b). This measure takes into consideration not only the number of pores, but also the pore size. Interspecific differences in the porosity of planktonic foraminifera are quite pronounced (Murray 1991b). These differences become especially apparent when viewed from a latitudinal perspective (i.e., temperature). Thus, we find that test porosity is low in higher latitudes and high in low latitudes, which roughly correlates with temperature differences. For example, Neogloboquadrina pachyderma, which is a high-latitude form (i.e., subpolar), has a mean test porosity of about 2%, while Globigerinoides sacculifer, which is abundant in the Indonesian waters (i.e., tropical), has a mean porosity of 18% (Murray 1991b). However, there are also significant intraspecific differences in test porosity which are related mainly to ontogeny (i.e., history of development). Unfortunately, only a few life history studies on planktonic or benthic foraminifera have been conducted thus far (Brummer et al. 1987), and none have yet been attempted in Indonesia. Intraspecific differences in test porosity have also been used to differentiate between cold-water and warm-water pelagic species (Be 1968,1977; Be et al. 1973). In their study of the Indian Ocean foram Orbulina universa, Be et al. (1973) demonstrated that populations from low latitudes (i.e., the equatorial Indian Ocean) had a significantly higher porosity than foraminifera in higher latitudes. O. universa has been reported from the Savu Sea (Adisaputra 1989), Segara Anakan, South Java (Subardi et al. 1989) as well as from Ambon Bay (Hermanto and Suhartati 1989), however, no information on its test morphology is available. An explanation for intraspecific differences in test porosity has so far been very elusive; thus while there are demonstrable effects from such factors as temperature, light, salinity, nutrient supply and seawater density, the development of test porosity and its functional role are still a mystery (Be 1977; Hemleben et al. 1985; Hemleben etal. 1987).
Test Coiling
A number of foraminiferal families (planktonic and benthic) exhibit pronounced test coiling. A trochospiral condition exists when foram test chambers are spirally coiled, evolute (i.e., unfolded and where all whorls are visible) on one side and involute (i.e., tightly coiled with only the final whorl visible) on the opposite side (e.g., Trochamminidae, Globorotaliidae, Eponididae, Nonionidae). The coiling of the test can be either dextral (i.e., clockwise, coiling from left to right) or sinistral (i.e., anticlockwise, coiling from right to left). Brummer and Kroon (1988) have demonstrated experimentally that coiling in foraminiferal tests is to a large extent genetically determined. However, this does not explain why a particular species would have both dextral and sinistral growth-forms; the functional significance remains elusive. Of particular interest, however, is the fact that some species have distinct geographic populations whose members are predominantly trochospiral in one direction only, while a population in another geographic area exhibits an opposite coiling direction. The cold-water planktonic foraminifera Neogloboquadrina pachyderma has become a classical example of this phenomenon (Arikawa 1983). N. pachyderma is distributed widely throughout the polar and subpolar regions. The polar and subpolar populations are at present most likely reproductively isolated from one another. In cold polar regions, coiling in N. pachyderma is generally sinistral, while in the slightly warmer subpolar regions the coiling is dextral. One is tempted to evoke temperature as the obvious controlling factor, and attempt to use this morphological feature in paleoclimatic analyses. However, Murray (1991b) suggested a cautious approach to the application of coiling ratios in paleoecological or biostratigraphic studies.
THE USE OF STABLE ISOTOPES
With advances in modern technology, new methods were developed that allow us to extract a wealth of stored environmental information from the foraminiferan tests, and other CaCO3-secreting organisms. The two techniques that have become indispensable to paleontologists, in their reconstructions of paleoclimates, are the analyses of stable (i.e., not radioactive) oxygen (8 O) and carbon (8 C) isotopes in calcareous skeletal material. There has been a recent surge in the use of other elements, such as nitrogen (815N), sulfur (834S), strontium and calcium ratios (Sr/Ca), boron (8 B) and hydrogen ( H and D - deuterium). The use of stable isotope analysis to solve various biogeochemical problems has increased dramatically in the past decade, mainly because stable isotope data are now used by both physical scientists (e.g., oceanographers, geologists) (Peterson and Fry 1987; Salomons and Mook 1987; Wu and Berger 1991; Beck et al. 1992; Lehman and Keigwin 1992; Farquhar et al. 1993; Plank and Langmuir 1993; Spivack et al. 1993) as well as by biologists in purely process-oriented biological or ecological studies (e.g., ontogeny, food-web analysis) (Eichler 1966; Weber 1974; Land and Lang 1975; Black and Bender 1976; Goreau 1977; Estep and Dabrowski 1980; Schroeder 1983; Peterson et al. 1985; Ehleringer et al. 1986; Rounick and Winterbourn 1986; Muscatine et al. 1989; Monteiro et al. 1991; Durako and Hall 1992; Murphy and Kremer 1992; Cerling et al. 1993). Stable isotope techniques are possible because each element has at least two stable isotopes (each with slightly different physicochemical properties), one of which is always present in greater abundance than the other. As the elements cycle through the biosphere, they undergo numerous chemical reactions that alter the isotopic composition (i.e., fractionation) of the elements in a predictable way (Peterson and Fry 1987), and these differences can now be measured with great accuracy.
Stable Oxygen Isotopes
Oxygen (O2) concentrations in seawater have long been used by physical oceanographers to trace the origins of various oceanic water masses, and together with salinity to determine their mixing rates (Broecker 1974). Oxygen became a powerful oceanographic tool when it was found that the two stable isotopes (i.e., heavy 18O and light16O) that comprise the oxygen element have subtly-differing physicochemical properties. Water molecules bearing 18O (i.e., heavier isotope) evaporate much slower from the sea, but condense faster than water molecules containing 16O; thus a "fractionation" of oxygen occurs as the element cycles through the atmosphere. This has a great influence on the isotopic composition of rainwater as well as the water masses from which the water initially evaporates. Oxygen-18 is present in seawater in much smaller quantities than 16O; in fact, 99.8% of all water molecules in the oceans bear 16O and only 0.2% bear 18O. Various water masses in the world's oceans are characterized by specific 18O/ 16O ratios, and a generalization can be made that surface waters in high latitudes contain lower 18O content than those in the equatorial regions (Broecker 1974). In other words, warm tropical water, because of its excess evaporation, becomes enriched in 18O, while high-latitude cold water becomes enriched in 16O. Another important feature is that the 18O/ 16O ratio is also salinity-dependent. High-latitude water masses characterized by low 18O content also have low salinities (Whitman and Berger 1992), while high-salinity equatorial water masses are enriched with 18O.
The differences in the physico-chemical properties of the two oxygen isotopes have provided a powerful tool for extracting paleoenvironmental information from the fossil record ever since the technique was first introduced (Emiliani 1955). Oxygen isotope analysis of the calcareous remains of foraminifera, corals and other calcifying organisms (e.g., Halimeda) has been applied widely in environmental and ecological studies (Pätzold 1984; Margosian et al. 1987; Siegrist and Randall 1989; Klein et al. 1992; Oba et al. 1992). All calcifying aquatic organisms must incorporate stable isotopes into their CaCO3 skeletons during the calcification process, the source of the oxygen being the CO2 as either HCO3- or CO3-. Among the various marine calcifying organisms, the isotopic ratio in foraminiferal and coral CaCO3is usually close to isotopic equilibrium with seawater (Urey 1947; Emiliani 1955, 1978; Wefer and Berger 1991). Note that in foraminifera and corals, some degree of δ18O fractionation occurs. One explanation for generally low δ18O values (relative to equilibrium) in scleractinian skeletons is that the HCO3- used during the calcification may be in isotopic disequilibrium with seawater (McConnaughey 1989a,b), Spero and Lea (1993), working with Globigerinoides sacculifer under laboratory conditions, showed that shell δ18O values vary as a function of symbiont photosynthesis, but do not vary with ontogeny. Nonetheless, the δ18O technique is very useful, since any changes in the ambient (i.e., seawater) isotopic composition will be reflected in the isotopic composition of the CaCO3 deposited at the time. Thus, the skeletons (e.g., foraminiferan tests, coral coralla, bivalve shells, cephalopods shells, etc.) in fact become recorders of the environmental (e.g., seasonal temperatures) as well as physiological changes that occur during the lifetime of the organism (Romanek et al. 1987). Based on samples recovered from deep-sea sediment traps set in the Sargasso Sea, Deuser et al. (1981) were able to demonstrate that in Globigerinoides ruber, which is the most frequently used species for isotopic determinations in Indonesian studies (Ganssen et al. 1989; Gayet et al. 1990; Barmawidjaja et al. 1993), deposition of CaCO3 occurred close to isotopic equilibrium with surface seawater. Experimental laboratory studies with Globigerinoides sacculifer and Orbulina universa demonstrated that CaCO3 deposition indeed occurs under isotopic equilibrium with seawater, and that changes in oxygen isotopic values are directly related to temperature, provided all other variables remain constant (Erez and Luz 1983; Bouvier-Soumagnac and Duplessy 1985). Thus these species are useful biomarkers. There are, however, exceptions to the equilibrium model, as, for example, in Globigerina bulloides, an indicator of upwelling conditions in the Banda Sea, whose tests in the Sargasso Sea (North Atlantic), are known to be deposited in isotopic disequilibrium with surface waters (Deuser et al. 1985). Thus analysis of oxygen isotopes in this species would be of limited value without additional data.
The 18O/16O isotopic ratio in the CaCO3 skeletons of modern foraminifera is primarily dependent on the ambient seawater temperatures (Urey 1947), salinity (Craig 1965; Tan and Strain 1980; Whitman and Berger 1992) and the isotopic composition (i.e., 18O/ 16O ratio) of seawater from which the carbonate is precipitated (Patzold 1984; Romanek 1987; Whitman and Berger 1992). The analysis of 18O/16O in CaCO3 skeletal material is somewhat complicated, because the temperature effect may be masked either by oxygen isotopic changes due to variations in physiological activity, especially in organisms associated with photoendosymbionts (e.g., foraminifera, corals, giant clams), or by salinity differences. However, under constant salinity and 18O/16O ratio of seawater (i.e., most oceanic coral reef environments), temperature has an overriding influence on the incorporation of oxygen isotopes into the foraminiferal tests, and other CaCO3 skeletons. In general, as seawater temperatures increase, the 18O/ 16O ratio in the carbonate decreases correspondingly and vice versa. This is very useful information, since using the isotopic ratios, from foraminiferan tests, for example, we can calculate the seawater temperatures under which the CaCO3 was deposited. Thus, the tests of Globigerinoides ruber, which has a wide geographic distribution, will contain more 18O if the formation of the test occurred in a cold-water environment (e.g., upwelling area, or winter months), while the tests will have lower 18O content if the tests were formed in a warm-water environment. Indeed, this relationship was observed in plankton tow studies from Bermuda (Williams et al. 1981).
However, great caution is needed to interpret oxygen isotopic data in organisms such as planktonic foraminifera, since throughout their different life-cycle stages they require different environmental conditions (i.e., depth-temperature related), and thus the continuous deposition of CaCO3 could proceed under very different temperature regimes, which is reflected in the isotopic signature. For example, most juvenile planktonic foraminifera (especially symbiotic species) mature in the warmer surface waters (0-30 m), and upon reaching sexual maturation they migrate into deeper-cooler waters just prior to gametogenesis (Murray 1991b). It is now believed that they migrate down to the top of the seasonal thermocline and DCM (H. Spero, pers. comm.). As expected, isotopic ratios in the tests of these species will reflect the extent of the vertical migration, since it is registered in the CaCO3 by 18O enrichment. Obviously, knowledge of their ontogeny is essential to unravel these life-cycle-related complexities in the isotope record (Murray 1991b).
Ice Volume Effect
Another factor that needs to be mentioned when discussing the δ18O technique is the magnitude of the ice effect (Emiliani 1955, 1966). This effect is quite considerable, since large volumes of water are moved from ice to sea during the glacial/interglacial cycle, causing large-scale sea level fluctuations (Milliman and Emery 1968). The maximum sea-level drop during the last glacial period in regions outside the archipelago have been estimated between 120-130 m below present-day sea level (Milliman and Emery 1968; Chappel and Shackelton 1986; Fairbanks 1989). Recent studies on sea-level fluctuations in the Indonesian Archipelago indicate that during the last glacial maximum, the lowest sea level stand was somewhere between 148(±16) m and 106(±11) m below present-day sea level (Hantoro 1992). This implies that during the last glacial maximum, about 3.5% of ocean volume was stored in polar ice caps and continental glaciers. According to Whitman and Berger (1992), δ18O values of polar ice were between -38%o and -34%o (i.e., ice is greatly depleted in 18O). During glaciation, 18O preferentially stays in oceans (it is fractionated), thus increasing the seawater 18O/16O ratio (δ18O increases), and as expected, the foraminifera that happened to live during this period will have tests that are correspondingly enriched in the heavier isotope (i.e., δ 18O increase). 18O During deglaciation, when the glaciers release large volumes of water, the 8values decrease correspondingly. The difference in δ18O between the glacial and interglacial periods (i.e., ice effect) in deep-sea sediments is 1.2%o, which means that for every 10 m change in sea level the δ18O values change by 0.1 %o (Whitman and Berger 1992). Thus the δ18O sediment record has also been used as an important sea-level marker. Foraminifera have stored all this information in their CaCO3 test.
Measurement
All stable isotopic ratios are measured with a mass spectrometer (Schneider and Jones 1992), and the results are by convention presented in delta (8) values, which are expressed as deviations per mill (i.e., %o) relative to an international standard PDB (Peedee Belemnite CaCO3). The delta values are calculated as:
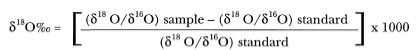
The δ values are thus ratios of heavy and light isotopes in the sample, compared to a known standard. The standard used depends on the analysis being done. For example, in δ18O analysis of foraminiferan tests or coral skeletons (i.e., calcite or aragonite) the standard is PDB. It is useful to remember that any increase in 'δ' value implies a corresponding increase in the heavier isotope. The δ18O is now a widely used technique in paleontology, since δ 18O isotope analyses of deep-sea cores containing calcareous organisms can reveal information on seawater temperatures at the time when the animal was alive and depositing the skeleton, which may have been millions of years ago. One equation commonly used to convert the δ18O values to temperatures is the paleotemperature equation developed by Epstein and Mayeda (1953):
![]()
where δ 18Oc and δ18Ow refer to shell and water samples, respectively. However, the isotopic composition of seawater varies with salinity, which is a function of evaporation, precipitation, freezing, mixing, and fresh-water runoff. To circumvent some of these problems, reconstruction of paleotemperatures has relied heavily on the use of foraminiferal shells from deep-sea sediments. One limitation of the present δ18O technique is that the temperature interpretation of foraminiferal isotopic data extracted from deep-sea sediments is based largely on our knowledge of modern foram physiology, and their response to temperature changes under laboratory conditions. However, new techniques are being developed that may circumvent some of these problems. One of the most promising is the strontium/calcium ratio technique (Beck et al. 1992), since the incorporation of the two elements into the CaCO3 skeletons (i.e., aragonite) is a function of temperature, but only a weak function of salinity (Beck et al. 1992). Using this technique, Beck et al. (1992) suggested that about 10,200 years ago the average sea surface temperatures in the southwestern Pacific Ocean were about 5°C cooler than they are today, and that the tropical belt was much narrower. Sr/Ca studies using massive skeletons of Pontes spp. have apparently been conducted in Indonesia; however, the results are not available.
Use of δ18O in Ontogenic Studies
To digress a bit, we should point out that oxygen isotopes can be very useful in tracing life history (i.e., ontogenic) changes in habitat depths of other organisms which may be difficult to observe in their natural habitats. A prime example is Nautilus pompilius (Cephalopoda), a deep-water, reef-associated cephalopod which is frequently sighted in deeper regions (>50 m) of the reef slope on many reefs in the Pacific region (fig. 9.15).
The population demography of Nautilus in Indonesia is still a mystery, yet its shells are sold in great quantities in almost all souvenir and handicraft stores in the country, even though it is a fully protected species. The fossil record of the extant Nautilus dates to the Oligocene; however, the Subclass Nautiloidae (i.e., cephalopods with calcareous external multichambered siphunculate shells) first appeared in the late Cambrian, and their basic body plan has not been altered since the Ordovician (O'Dor et al. 1993). Nautilus javanus from the late Miocene is the only fossil cephalopod described from Java (van Bemmelen 1949). There are a number of extant species, the most common being Nautilus pompilius, which is a true living fossil (Saunders 1987). This deep-water invertebrate is known on occasions to visit the deeper parts of the lower reef slopes, but the adults usually stay at depths below 100 m, and none have so far been captured at depths of less than 50 m (Obaetal. 1992).
The use of δ18O was instrumental in unraveling the mysteries of its ontogenic changes in habitat depths. We should also point out that successful hatching of aquarium-held Nautilus belauensis has greatly expanded our understanding of its ontogeny (Carlson et al. 1993). It appears that Nautilus has an exceptionally long incubation period lasting about 12 months. According to Oba et al. (1992), incubation takes place in the shallowest depths of its overall depth distribution, which may, however, vary significantly between different regions depending on other environmental conditions. Oba et al. (1992) were able to demonstrate that nautili in the Philippines (i.e., Tañon Strait) hatch at depths between 100-80 m (24°-28°C). Based on the δ18O values in nautili shells, it was demonstrated that once hatched, the juvenile N. pompilius live at the hatching depth until the deposition of the twelfth septum. The juveniles then move into deeper water where temperatures are about 17°C, and can be as low as 13°C, which is reflected in the sharp increase in δ 18O values, from about -0.3%o (or lower) to values roughly between 0.6%o to 1.2%o (Oba et al. 1992). The use of δ18O values was also instrumental in determining that Nautilus in Fiji inhabit considerably deeper waters, which was reflected in enrichment in δ 18O in both juvenile (range 0.1 %o to 0.5%o) and adult (range 1.4%o to 2.0%o) nautili. Thus hatchlings and juveniles are found at depths between about 300 to 350 m, while adults live at depths between 450 to 550 m (Oba et al. 1992).
Figure 9.15. A true living fossil, Nautilus pompilius is a deep-water reef-associated cephalopod that occurs throughout the Pacific from Fiji to Indonesia.
Photo courtesy of R. Steene.
The usefulness of the δ 18O technique was further demonstrated by analyzing the isotopic composition of the last septum of mature adults. Micro-sampling taken from the last septum showed regular fluctuations in δ18O values of about 0.3%o to 0.5%o, roughly corresponding to differences in 1°-2°C. These regular δ18O fluctuations correspond to daily vertical migrations of the species. This has been corroborated with telemetric studies at the University of Papua New Guinea, Motupore Island Research Station (O'Dor et al. 1993).. Vertical movements of Nautilus along the reef face are mostly crepuscular, ascending at dusk to shallower reefal waters in search of food and descending into the deeper, cooler waters at dawn, presumably to conserve energy (O'Dor et al. 1993).
Recent isotopic studies, as well as successful hatchings of nautili at the Waikiki Aquarium (Carlson et al. 1993), have provided important clues to the ontogeny of this living fossil. The requirement for warm water for incubation seems to have answered one of the major questions of Nautilus distribution in the tropics (Carlson et al. 1993). Thus, while the species is obviously temperate in character, since it prefers cooler deeper waters, they need the tropical warm water for incubation. Thus, their reproductive strategy has restricted their distribution to the tropical oceans. It is believed that Nautilus has only a limited ability to swim between islands (Saunders 1987), and thus the absence of a planktonic larval stage during its ontogeny greatly reduces its dispersal ability, and partly explains its rather narrow distributional range.
STABLE CARBON ISOTOPES
Perhaps of greater value in recent paleontological investigations has been the use of stable carbon isotopes (i.e., 12C and 13C). Carbon-12 is much more abundant than 13C, representing 98.89% of the carbon pool, while 13C accounts for the additional 1.11%. Studies of 12C/13C (i.e., δ13C derived as for δ18O) ratios in animal skeletons (especially foraminifera) have provided valuable information on past carbon cycles, and most importantly on the productivity of the paleoceans. In δ13C determinations the international standard is Belemnitella americana from the Cretaceous PeeDee Formation (PDB). Unlike the oxygen isotopic ratios, the δ13C is not temperature-dependent. This means that the δ13C value in shells of various calcifying organisms, or coral skeletons, is very close to the ambient values in which the CaCO3 was precipitated, and is thus useful in assessing productivity (i.e., photosynthesis) of pale-oceans at the time of CaCO3 deposition. Indeed, high δ13C values in foraminifera tests have been correlated with high primary productivity of the water masses in which the foraminifera deposited their tests. The δ13C enrichment occurs because during phytoplankton blooms, or in regions with intense primary productivity, phytoplankton preferentially take up 12C, which leads to a substantial enrichment of C in the ΣCO2 of surface waters (Ganssen et al. 1989). During photosynthesis 12C atoms are fixed 1.02 times more rapidly than 13C atoms. Since δ13C in CaCO3 skeletons reflect ambient δ13C levels, foraminifera from regions of high primary productivity will have tests usually enriched in δ13C (Ganssen et al. 1989).
Carbon isotopes have also been useful in ontological studies of planktonic symbiotic foraminifera. Tests of juvenile symbiotic planktonic foraminifera have been shown to be depleted in 13C (i.e., lower δ13C) when compared to larger individuals. The isotopic difference is a result of vital effect related to the photosynthetic activity of the photoendosymbionts (mostly dinoflagellates and chrysophytes).
Juvenile foraminifera are usually found in the upper layers of the euphotic zone until maturity, and, as a result, photosynthetic activity of the endosymbionts is high (Spero and Parker 1985), thus producing ample quantities of isotopically light (i.e., C depleted) carbon pool which is used by the juvenile foram in CaCO3 deposition. As a result, the tests of all small (i.e., juvenile) symbiotic foraminifera are δ13C depleted.
Differences in δ13C values and morphological variations in test architecture are powerful tools to distinguish different foraminiferan sub-populations. Perhaps one of the best examples is that of Globorotalia truncatulinoides, whose subantarctic and subtropical populations in the southern Indian Ocean differ in their δ13C signatures and test morphology (Healy-Williams et al. 1985).
PALEOCEANOGRAPHY AND PALEOCLIMATES
Stable isotopes have become important portholes into the distant past, and have provided new insight into the paleoclimatic and paleoceanographic conditions during the last glacial period in the Indonesian Archipelago. It is well-established that, during the last glacial maximum, sea surface temperatures in most equatorial oceanic regions dropped by about 2°-3°C, with a corresponding equatorward displacement of polar fronts (Seibold and Berger 1982). Surface waters of many subtropical and tropical coastal environments (e.g., eastern Indonesia) may have been additionally cooled by the intensification of coastal upwelling. As expected, δ18O values of planktonic foraminifera (recovered by sediment cores) in many regions are enriched in 18O, reflecting the general cooling of the oceans. Note that the temperature enrichment is in addition to the ice volume effect that globally accounts for about 1.2%o of the difference between the last glacial maximum and the Holocene high sea level stand (Seibold and Berger 1982).
Isotopic analysis of deep-sea foraminiferal sediments has provided important clues concerning paleoclimates and paleoceanography since the last glacial. New techniques using foraminiferal boron isotope (δ11 B) extended the record to the early Miocene (about 21 Ma). Spivack et al. (1993), using a foraminiferal boron isotope as a proxy for surface ocean pH over the past 21 Ma, demonstrated that the surface seawater pH from the early to middle Miocene was between 7.4 to 7.5 pH. This measurement was made possible because the isotopic composition of boron in foraminiferal tests is pH-dependent. Since the pH of surface waters is a sensitive measure of its alkalinity and total inorganic carbon concentrations, Spivack et.al. (1993) concluded that total carbon and alkalinity 21 Ma were 0.6 times higher than present-day values, indicating that the CO2 concentrations of the Miocene atmosphere were much higher than at present. By late Miocene (7.5 Ma B.P.) surface seawater pH values reached 8.2 and remained so up to the present. The Miocene was a period of extensive coral reef development with a global climate that was much wetter and warmer. It seems that lower pH values (higher atmospheric CO2 concentrations) did not have a detrimental effect on coral reef development at the time. Recent reviews on the effects of global climate change on coral reef ecosystems have, however, suggested that, with an increase in sea surface temperatures and atmospheric CO2 concentrations, a decrease in CaCO3 saturation state may be expected (Smith and Buddemeier 1992). Smith and Buddemeier (1992) further suggested that decreased rates of calcification associated with the altered CaCO3 saturation state will reduce the ability of coral reefs to keep up with the expected sea-level rise. It is interesting to point out that low sea-surface pH values (i.e., lower CaCO3 saturation state) from early to middle Miocene were associated with rising sea levels; however, since the end of the middle Miocene, the steadily rising sea-surface pH values (Spivack et al. 1993) were accompanied by a major glacio-eustatic lowering of sea level, reaching one of the all-time lows (c. 220 m) at about 6.6 Ma B.P. (Hutchison 1989).
Three recent studies in Indonesia are profiled to illustrate the great value of stable isotope techniques as well as to review some exciting new information on the paleoclimate and paleoceanography of the region. In concert, these studies offer an incredible glimpse into the glacial past, and provide an interesting chronological sequence of events since the last glacial maximum. The following reconstruction of past events would not have been possible without the environmental record that was extracted from the tests of small foraminifera.
NORTHERN MOLUCCA SEA
The analysis of the Molucca Sea deep-sea sediment core (discussed earlier) revealed that δ18O values during the last glacial maximum were 0.8%o higher (18O enriched) than would be expected if ice volume effect was the dominate factor controlling the δ18O values in planktonic foraminifera (Barmawidjaja 1993). Based on their palynological, planktonic foraminifera and δ 18O results, Barmawidjaja (1993) concluded that the 18O enrichment during the last glacial maximum in the Molucca Sea was mainly a function of higher salinities, and not temperature. Higher salinities in the surface waters were brought about by increased aridity in the region. They hypothesized that increased aridity during the last glacial resulted in higher evaporation rates, relative to precipitation, which decreased the δ18O values (i.e., 18O enrichment). What caused the increased aridity during the glacial maximum remains equivocal; however, Barmawidjaja et al. (1993) suggested that the causative mechanism is most likely a combination of two climatic factors driven by the asymmetrical distribution of glacial ice cover. While they acknowledge that the Inter-Tropical Convergence Zone (ITZC) did not oscillate as widely (i.e., a reduction in the northern extent) as it does today, it was not the main cause for the increased aridity. To explain this phenomenon, Barmawidjaja et al. (1993) proposed that the most plausible explanation for the increased aridity during the last glacial maximum lies in the possible disruption of the El Niño-Southern Oscillation (ENSO) system. It seems that during deglaciation, the ENSO system resumed its course, and the climate in the eastern regions of the archipelago became progressively wetter (fig. 9.16).
The appreciable and rapid reduction in δ18O values from about 14 ka to 8 ka (fig. 9.16) corresponds to the last deglaciation, and the rapid melting of the continental glaciers and polar ice caps. These results agree well with those of Ganssen et al. (1989), who arrived at similar conclusions.
The northern Molucca Sea deep-sea core (Barmawidjaja et al. 1993) provided a wealth of information on the climatic conditions that existed in the region during the last glacial maximum. One of their most significant findings, from the coral reef perspective, is that surface seawater temperatures in the northern Molucca Sea during the last glacial maximum (i.e., 18,000 ka) were at most only 1°C cooler than at present, and that the 18O enrichment was mainly salinity-driven. This suggests that glacial sea surface temperatures were basically within the optimal range for corals and coral reef development, providing the first conclusive evidence that temperature fluctuations during the Plio-Pleistocene were not a significant factor affecting coral distribution and coral reef development in the archipelago.
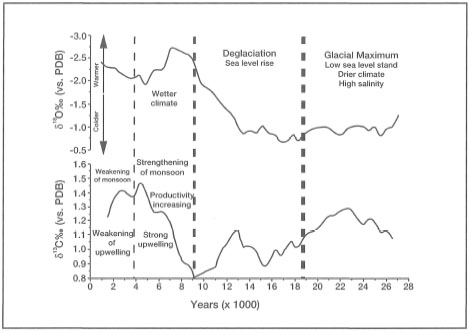
Figure 9.16. Oxygen (δ18O) and carbon (δ13C) isotope record for Globigerinoides ruber from the northwestern Moluccas Sea plotted against time. Original data modified by smoothing procedure to enhance long-term patterns (i.e., glacial - interglacial differences). Time scale is refined chronology based on AMS radiocarbon (14C) dates of monospecific samples of Globorotalia menardii-tumida.
Source: Barmawidjaja et al. 1993 and Ganssen et al. 1989.
MAKASSAR STRAIT
Using foraminiferal biomarkers (species assemblages), and δ18O of foraminiferan tests from sediment cores taken in the Makassar Strait, Gayet et al. (1990) were able to reconstruct paleoclimatic conditions in the western region of the archipelago during the past 15.5 ka. The results provide additional support to earlier studies demonstrating that the last deglaciation was interrupted by the Younger Drays event (Fairbanks 1989), which is reflected in δ 18O enrichment of planktonic foraminifera (Ganssen et al. 1989; Linsley and Thunell 1990). Following the glacial maximum at about 18.8 ka, the abundance of planktonic foraminifera as well as surface water productivity in the Makassar Strait were below present-day levels. Most of the sediments in the Makassar Strait at the time were allochthonous, originating from Kalimantan via the Mahakam River discharge. The δ18O values of planktonic foraminiferal tests showed that from 15.5 ka to 13.3 ka there was a sharp discontinuity in the δ18O values from -0.5 to -1.1 %o, corresponding with rapid meltdown of the polar ice caps. These results have been corroborated by a more recent study in the northern Molucca Sea (see fig. 9.16) (Barmawidjaja et al. 1993). From 13.3 ka to about 10 ka melting of the polar ice caps was slowed, and more stable conditions prevailed. Increased productivity in the surface waters (i.e., higher abundance of planktonic foraminifera) was most likely associated with significant weakening of the water column stratification, which allowed greater mixing between surface and deep water masses. Greater vertical mixing facilitated by the weakening of the water column stratification may have resulted in higher oxygen concentrations of the deep water masses, which increased from about 5.0 mg.l-1 to about 5.7 mg.l-1 by 13.3 ka (Gayet et al. 1990). Since about 8.2 ka, circulation in the Makassar Strait seemed to have slowed down, mainly as a result of the reestablishment of a relatively strong stratification during the Holocene. As a result, there has been a measurable reduction of oxygen in the bottom water masses (Gayet et al. 1990).
BANDA SEA
During the Snellius-II Expedition, an interesting study was conducted in the Banda Sea region to reconstruct the conditions which existed during the last deglacial period between 15 ka to 8.2 ka. Using δ18O techniques, Ganssen et al. (1989) were able to show that the last deglaciation did not lead to lower sea surface temperatures in the Banda Sea region, but that it had a considerable influence on the regional weather patterns. As expected, climatic conditions and oceanic circulation patterns 10,000 years ago were much different. Palynology of deep-sea sediment cores revealed that the climate in eastern regions of the archipelago during deglaciation was much wetter, while the upwelling intensity increased significantly, resulting in increased productivity of surface waters, reflected in higher δ13C values (fig. 9.16). Their δ18O record of the core clearly demonstrated that deglaciation was interrupted by the Younger Drays event. There appears to be a significant difference in the sensitivity of the marine and terrestrial systems to climatic changes. The response of terrestrial flora (i.e., increase in spore/pollen ratio) to the intensification of the monsoonal system (i.e., wetter climate) began about 1300 years earlier than the corresponding increase in primary productivity (higher δ13C - see fig. 9.16) of surface waters (Ganssen et al. 1989). However, as illustrated in figure 9.16, the increase in primary productivity since about 9 ka has been remarkable.
The driving mechanism behind the high productivity of the surface waters in the eastern regions of the archipelago is upwelling, driven by the monsoonal system. What we learned from the tests of planktonic foraminifera is that the disruption of the monsoonal system will ultimately lead to a reduction in marine productivity. More importantly, however, is that weakening of the present monsoonal system will result in rapid climatic changes in the eastern regions of the archipelago. This is clearly evident from the recent ENSO events, which were associated with long periods of drought, conditions that may have existed during the last glacial. The anticipated climate change (Houghton et al. 1990) is a serious concern for Indonesia, since the most likely scenario will be a major disruption (i.e., weakening) of the monsoonal system. The paleoclimatic information extracted from the deep-sea sediment cores indicate that, if this occurs, a change to more arid conditions can be expected, perhaps within the life span of the next generation.
CONTRIBUTION TO MARINE SEDIMENTS
Large areas of the ocean floor are covered by pelagic biogenic sediments, consisting mainly of calcite tests of holoplanktonic foraminiferans, such as the Superfamily Globigerinacea (e.g., Globorotalia, Globigerina, Globigerinoides, Globigerinella). Globigerinoides are a cosmopolitan group which has, however, not been studied in Indonesian waters even though they are a significant component of the zooplankton community (fig. 9.17).
After millions of years of continual deposition, the calcareous foram tests constitute an important component (>75%) of deep oceanic sediments. Other ingredients of deep oceanic sediments are the remains of calcareous holoplankton, such as the calcitic coccoliths (Prymnesiophyta; usually < 10 μm in diameter) or aragonite-secreting pteropods (i.e., planktonic gastropods), as well as chitinous remains of copepods (Crustacea), tintinnids (Tintinnidae) and other zooplankton (van Waveren 1989a,b, 1993).
We should point out, however, that in other oceanic regions, notably in the higher latitudes as well as in the equatorial east and central Pacific, deep oceanic sediments consist mainly of siliceous (amorphous hydrated silica or opal SiO2nH2O) remains of diatom (Bacillariophyta) frustules or radiolarian (Radiolaria) skeletons (fig. 9.18).
In areas of high productivity, such as the Banda Sea, diatoms can be so productive that their frustules form diatom oozes (Situmorang 1989). In ocean basins where deposition occurs above the lysocline (i.e., depth at which dissolution of calcareous material takes place), foraminiferal tests may form deposits hundreds of metres thick, which are often referred to as "globigerina ooze". In the Banda Sea, foraminiferal ooze is the dominant biogenic deposit, and has been classified according to composition and texture (fig. 9.19). One type consists mostly of tests of pelagic foraminifera Globorotalia spp. and Globigerina spp. and is of a loose sandy character, while a more clayey foram ooze consists of foraminifera in a sticky material (Situmorang 1989).
Foraminiferal ooze in fact covers about one-half of the world's deep-sea floor, and thus is the most widespread deposit on the planet. This is especially true along submarine ridges, where depositional environments are generally above the lysocline depth, which varies from sea to sea. Lysocline depth, and thus the fate of foraminiferal tests and other calcareous fragments, is controlled by a number of factors, but mainly by temperature and pressure (Be 1977). Calcium carbonate becomes progressively more soluble with increasing depth (i.e., increase in pressure) and decreasing temperature. However, the dissolution susceptibility of foraminiferan tests is also highly species-specific, since it is also related to their general morphology. The tests of spinose planktonic foraminifera, such as Globigeri noides ruber, G. sacculifer and Globigerinella aequilateralis, undergo rapid dissolution as they sink to the bottom, which has a significant effect on the composition of bottom sediments, especially in areas below the lysocline (Troelstra et al. 1989). We should point out, however, that complete dissolution of the foraminiferan tests (and all other calcareous material) takes place mostly on the seabed. More resistant to dissolution is a group of non-spinose foraminifera with thicker tests, such as Neogloboquadrina dutertrei, Pulleniatina obliquiloculata and Globorotalia tumida, which tend to dominate bottom sediments (Troelstra et al. 1989).
Figure 9.17. Percentage of ocean floor covered by pelagic biogenic sediments.
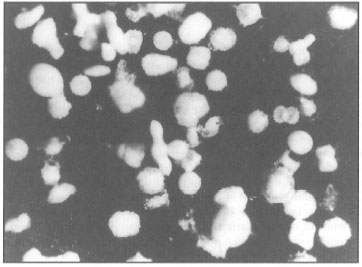
Figure 9.18. Radiolarian ooze from the Argo Abyssal Plain, depth 5375 m. Leg G.6-2 of the Snellius-II Expedition.
Photo courtesy of M. K. Adisaputra, Pusat Pengembangan Geologi Kelautan -LIPI, Bandung.

Figure 9.19. Globigerina ooze dominated by planktonic foraminifera Globorotalia menardii. Location: Lombok Ridge, depth 3150 m, Leg G.6-4 Snellius-II Expedition.
Photo courtesy of M. K. Adisaputra, Pusat Pengembangan Geologi Kelautan -LIPI, Bandung.
In the Indonesian Archipelago, the depth of the lysocline in the various basins varies from region to region, depending on the local hydrological conditions. In the Weber Deep, the lysocline is located at a depth of 1700 m, and is characterized by rapid dissolution and fragmentation of foraminiferan tests as well as a significant increase in the abundance of radiolarians. Since radiolarian skeletons are siliceous, they are less prone to dissolution. In addition they may be covered by a thin organic layer providing additional protection. In the Seram Trench, the lysocline is located at a depth of 1100 m, which seems to be one of the shallowest in the archipelago. According to Troelstra et al. (1989), the shallower lysocline in the Seram Trench is associated with strong local upwelling processes, which bring up cooler and calcium-carbonate-undersaturated deep water masses that cause rapid dissolution of foraminiferan tests. The same fate awaits the pteropod (i.e., planktonic mollusc) shells, which are more prone to dissolution, since they consist of pure aragonite. Aragonite is much more soluble than calcite, which is the main constituent of foraminiferan tests. A number of recent studies have been conducted that allow a simple comparison of lysocline depths. Barmawidjaja et al. (1983), working in the northern Molucca Sea, just northwest of Halmahera, found that in an area dominated by the Mindanao Current, the lysocline was located below a depth of 3510 m. In the Sulu Sea, located to the northwest of the Sulawesi Sea, Linsley et al. (1985) reported lysocline at 3800 m, and Berger et al. (1976) reported a lysocline depth of 4100 m in the west Pacific. In the Lombok Basin (Indian Ocean), Adisaputra (1989) positioned the lysocline at a depth of 3157 m, and at 3300 m in the Savu Sea. In the Argo Abyssal Plain south of Nusa Tenggara, the lysocline was at a depth of 4200 m. Thus it seems that the shallowest lysoclines occur in regions of upwelling.
In general, deep oceanic water masses are undersaturated with respect to calcite and aragonite. Thus, at a certain depth (>5000 m), most, if not all, carbonate sediments disappear. Oceanographers have coined the termcm-2carbonate compensation depthcm-2 (CCD) to refer to the depth at which carbonate skeletal material of pelagic organisms contributes less than about 20% of the total sediment. The CCD in the Weber Deep is at a depth of 3300 m (Troelstra et al. 1989). The deepest CCD occurs in the Argo Abyssal Plain at a depth of 5400 m (Adisaputra 1989). As would be expected, the CCD depends on temperature and pressure; however, additional factors such as pelagic and benthic productivity are involved as well. In oceanic systems, sedimentation (i.e., accumulation of sediments) is mainly a function of surface water productivity; thus sedimentation can be used as a proxy indicator of productivity, but with great caution. Sediment traps are frequently employed by oceanographers to determine the downward flux of organic matter. In a study conducted in the Banda Sea, Ganssen et al. (1989) reported sedimentation rates ranging between 11.8 to 15.6 cm.ka-1. In other parts of the archipelago, sedimentation rates may be much higher, especially in areas influenced by major river systems. Eisma et al. (1989) estimated that based on present-day sedimentation rates of about 60 cm.ka-1 (c. 150 mg.cm-1.a-1), the Flores Basin will be filled in 10 million years. Based on δ13C values (i.e., -27.24 to -23.72) it seems that most of the sedimenting material is of autochthonous origins.
Since SiO2 is less susceptible to dissolution than carbonate, the glassy skeletons of diatoms and radiolarians are the major component of oceanic sediments below the CCD, and at depths greater than 5000 m. Diatoms also tend to dominate in higher latitudes, or in regions where upwelling promotes high primary productivity (e.g., west coast of Peru), while radiolarian ooze may dominate in low latitudes.
Surface productivity of radiolarians, and other planktonic microfossils, is very seasonal in the Banda Sea. Thus, during the non-upwelling period, productivity is very low and surface samples will seldom contain more than a few specimens, yet examination of the deep sediment cores will reveal that productivity during the upwelling period must be considerable. The concurrence of the surface abundance of planktonic microfossils and their benthic assemblages is high in non-upwelling regions year-round, while in areas under strong seasonal upwelling, the concurrence is low during the Northwest Monsoon and high during the Southeast Monsoon. Troelstra et al. (1989) identified over 300 species of radiolaria with some impressive abundance figures. For example, the flanks of the Weber Deep can contain as many as 400,000 specimens per gram of CaCO3-free sediment. These assemblages are dominated by Dictyocorne turncatum, Botryocytris scutum, Phorticium pylonium, Ommatartus tetrathalamus and Lithomellissa thoracites (Troelstra et al. 1989). A high abundance of radiolarians (100,000 to 300,000 specimens per gram of CaCO3-free sediment) on the flanks of the Weber Deep has been explained by van de Paverd and Björklund (1989) as being a result of a localized upwelling that affects primarily the flanks of the Weber Deep. Upwelling on the west flank of the Weber Deep may develop during the Northwest Monsoon as the surface waters are driven to the east, and are being replenished by deeper water along the west flank (van de Paverd and Björklund 1989). The system reverses during the Southeast Monsoon with localized upwelling along the eastern flank of the Weber Deep. However, the Weber Deep has recently been considered not to be influenced by upwelling (Troelstra et al. 1989). As would be expected, higher radiolarian abundance are found in the Seram Trench and on the continental slope of Irian Jaya where upwelling is more pronounced. These assemblages are dominated by Dimelissa apis, Phormacantha histrix, Pseudocobus obeliscus, Lophophaena variabilis and Arachnocorys circumtexta (Troelstra et al. 1989).