Chapter Four
INTRODUCTION
There have been many inspired literary works written in the past that touched upon the beauty and the intricacies of coral reefs. The diversity of types, shapes and sizes of Indonesian coral reefs, as well as the outstanding richness of the various plant and animal species that live in, on and above these unique biogenic structures, has fascinated and bewildered many early as well as present-day naturalists. While the beauty of tropical coral seas and a quest for adventure may have been the initial lures, the early history of coral reef science was sustained by the enigma of flourishing productivity in the midst of mostly oligotrophic (i.e., nutrient- and plankton-poor) tropical oceans (Lewis 1981).
While coral reefs in Indonesia have recently received considerable public, media and government scrutiny, their remoteness and often difficult access makes them generally inaccessible, and thus invisible, to the majority of the population. This may at first be surprising, considering the fact that the Indonesian coastal zone supports approximately 60% of Indonesia's 182 million people, and 75% of cities with populations in excess of 100,000, in addition to the 60,000 coastal villages (Soegiarto and Polunin 1981; Darus 1985; KLH 1992). Human interactions with coral reefs in Indonesia date to prehistoric times, as is evident from a few shell-middens in Sumatra, Sulawesi and Irian Jaya (Soegiarto and Polunin 1981; Whitten et al. 1987; Whitten et al. 1988). Presence of various artefacts, such as perforated shell discs from Java, Timor and Flores, ear ornaments from dugong teeth and turtle shell from Java and Bali, suggest that prehistoric coastal people throughout the Indonesian Archipelago had a close link with the sea (Soegiarto and Polunin 1981). Nonetheless, the vast majority of the Indonesian population is predominantly landward-oriented, even though this may not always have been the case with the Javanese (Sya'rani and Willoughby 1985). It appears that in the not too distant past, subsistence and commercial exploitation of marine resources, particularly of coral reefs, was conducted by a few seafaring groups such as the Bajaus, Buginese, Butonese, Mandarese, Makassarese and Madurese, while the more populous groups such as the Javanese and Balinese were agriculturists (Sya'rani and Willoughby 1985). Even in the eastern (i.e., oceanic) regions of the archipelago, great distinction is made between the coastal (i.e., fishermen) and the upland (i.e., farmers) people, with the fisherman being frequently looked down upon. The point being made is that in view of the predominantly landward orientation of the majority of the Indonesian population, conservation and protection of marine resources is a long-term endeavor. Rapid degradation of coastal ecosystems, such as coral reefs, is partly related to the land/water opposition, which is reflected in the general attitudes of people towards the sea and its ecosystems. For example, coral reefs are viewed by many coastal people as places with lots offish, but not necessarily as living systems which are the corner-stone of the fishery. The widespread use of explosives and poison in coral reef fisheries is a case in point. However, new programs by the government and non-governmental organizations (NGOs) are under way to address these illegal activities. It is evident that current perceptions about the nature of coastal ecosystems need to be changed, and this can be done successfully only through conservation programs where education is a key component.
Perhaps in the present context the articulate words of Sir Alfred Russel Wallace, the distinguished 19th-century naturalist, founder of modern biogeography and cofounder of the theory of evolution, combined with the artistic impressions of Roger Steene and Ron and Valerie Taylor, are most appropriate to introduce us into the magical realm of Indonesian coral reefs (colour plates 4.1 to 4.4).
Passing up the harbor, in appearance like a fine river, the clearness of the water afforded me one of the most astonishing and beautiful sights I have ever beheld. The bottom was absolutely hidden by a continuous series of corals, sponges, actiniae, and other marine productions, of magnificent dimensions, varied forms and brilliant colours…. It was a sight to gaze at for hours, and no description can do justice to its surprising beauty and interest. For once, the reality exceeded the most glowing accounts I had ever read of the wonders of a coral sea. There is perhaps no spot in the world richer in marine productions, corals, shells, and fishes than the harbor of Amboyna.—WALLACE 1869
Coral reefs are found throughout the tropical and subtropical regions of the Pacific, Indian and Atlantic Oceans. In the warm subtropical and tropical seas, significant coral reef development occurs in clear, and usually oligotrophic, waters at depths of up to 40 m (Verwey 1931a), and in the more turbid coastal waters where vigorous reef growth is restricted to depths of less than about 15 m. This general statement certainly typifies coral reefs globally, but within the Indonesian context, we must add that vigorous coral reef development also occurs in oceanic nutrient-rich regions under the influence of seasonal upwelling. Indeed, the long-awaited Indonesian coral reef data that are now starting to trickle into the international literature indicate that one of the most diverse coral reef regions in the world occurs in a major upwelling area of considerable global significance. The Flores, Molucca and Banda Seas have recently been identified as contemporary centres of scleractinian coral diversity (i.e., generic and species) (Best et al. 1989; Hoeksema this book; Wallace this book).
The Indonesian Archipelago is one of the most tectonically active and geologically complex regions on earth. The climate of the archipelago is driven by macro scale movements of air masses between the Asian and Australian continents, which have a pronounced effect on the surface circulation patterns of the Pacific and Indian Oceans. The general current patterns of the archipelagic seas are thus driven by large-scale atmospheric-oceanic interactions that are, however, regionally and locally modified by strong tidal regimes and geologic features of the archipelago (i.e., island arcs, atolls, barrier reefs, submarine ridges and volcanoes). This complex interplay of atmospheric, oceanographic and geologic processes has resulted in one of the most dynamic marine environments on the planet. Indeed, the environmental complexity associated with strong seasonality of monsoonal weather patterns, tidal regimes, volcanism and tectonic activity is being recognized as one of the primary factors responsible for the great diversity of coral reef biota. The complex interplay of abiotic and biotic factors has resulted in a marine environment characterized by high habitat diversity, which is ultimately responsible for the overall richness of the marine flora and fauna of the Indonesian Archipelago (Best and Boekschoten 1988). The physico-geochemical complexity of the archipelago is one of the key features that makes this region clearly distinct from the superficially similar volcanic arc system in the Caribbean (Best and Boekschoten 1988).
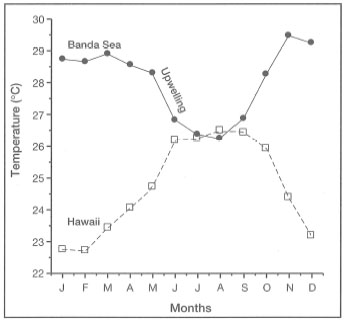
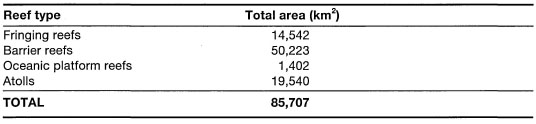
Figure 4.1. Comparison of the annual sea surface temperatures between the Banda Sea (Ambon and Haruku Islands) and the Pacific Ocean (i.e., Hawaii). Note that annual sea surface temperatures in the Banda Sea (i.e., upwelling region) are higher with slightly narrower (about 1 °C) seasonal fluctuations than sea surface temperatures around Hawaii.
An important characteristic of Indonesian coral reef environments, especially in the diverse eastern regions of the archipelago (e.g., Flores Sea, Banda Sea and Molucca Sea), is their relatively benign environmental state with regards to fluctuations in temperature or salinity. This is not to say that seasonal fluctuations of these variables, and other abiotic and biotic parameters, do not occur - they do, but they occur on a narrower scale than would be expected in regions influenced by strong seasonal upwelling (fig. 4.1).
Compared to the Great Barrier Reef Province, and other regions of the Indo-Pacific and southwestern North Atlantic, where coral reef ecosystems are frequently disrupted by severe natural disturbances (e.g., tropical cyclones, extreme low tides or low temperatures), the vast majority of Indonesian coral reefs are located in sheltered intra-archipelagic seas where severe physical perturbations are relatively rare. This is interesting, since it has been proposed that physical disturbances play a key role in the maintenance of high diversity and overall stability of coral reef ecosystems (i.e., the intermediate disturbance hypothesis) (Connell 1978). However, the main weakness of the intermediate disturbance hypothesis is that the magnitude and frequency of physical disturbances involved have not been quantified. While being removed from the severe effects of tropical cyclones, coral reefs in the various regions of the Indonesian Archipelago are nonetheless affected by tsunamis, volcanism, earthquakes as well as by intense, but short-duration, monsoonal storms. The effects of these natural disturbances on community structure and function of Indonesian coral reefs have yet to be documented. Ecological studies from the southwest Java Sea suggests that monsoons play an important role in structuring coral communities, at least in some regions of the archipelago (Suharsono 1992; 1994). However, Tomascik et al. (1996) have demonstrated that volcanic eruptions associated with andesitic lava flows can significantly enhance local coral reef diversity, by providing novel substrates for colonization. Whether high diversity of coral reef communities in the more sheltered regions of the archipelago (e.g., Togian Islands) is to a greater extent a function of biological interactions, such as predation, competition, and parasitism, remains to be determined. The diverse coral reef environments of the Indonesian Archipelago offer exciting possibilities to test a number of contemporary ecological hypotheses concerning the effects of natural disturbances on the structure and function of coral reefs.
CORAL REEFS UNDER THREAT
The diverse coral reef communities, so eloquently described by Sir Wallace (Wallace 1869) during his visit to Amboyna (i.e., present-day Ambon, Moluccas) in 1857, are becoming a rare sight in Ambon Bay. Most, if not all, coral communities in the vicinity of Ambon City have been destroyed mainly as a result of pollution, siltation and destructive fishing practices (modern and "traditional") (McManus and Wenno 1981; McManus 1988; Best et al. 1989). In the outer areas of Ambon Bay, one can still find reefs (Hualopu 1993), as described by Wallace, but their future survival is uncertain.
It took 138 years of "development" and "progress" to degrade a significant portion of the coral reef communities of Ambon Bay, situated in what has been a relatively remote location for the better part of Indonesia's modern history. The degradation is, however, not restricted to Ambon Bay. The once pristine reefs along the northeast coast of the islands are now marked by numerous bomb scars, a common sign of human presence even in the remote regions of this vast archipelago. The popular belief that traditional fishing practices, such as bamboo fish traps, are "environmentally friendly" is no longer acceptable in many coastal regions, where thousands of fish traps (instead of a few) are now being dragged over the reef daily, and many are left behind to rot (fig. 4.2). The "traditional" practice of camouflaging bamboo traps with coral boulders has resulted in widespread damage to coral reefs in many parts of the archipelago. This traditional practice is damaging, since live coral is taken from the productive and structurally important reef flat habitats. With more fishermen, and an easy access to mask and snorkel, more reef areas are now accessible to "traditional" exploitation, and thus, as the reef exploitation increases so does damage to the reefs.
Closer to major population centres on the more densely populated islands such as Java, with over 100 million inhabitants, degradation of coral reef communities, as well as physical destruction of the reefs themselves, has been much faster.

Figure 4.2. Abandoned fish trap on a deep (25 m) reef slope. Until the fish trap rots, it is a death chamber for reef fish. Without food the fish soon die, but their dead remains attract more fish, thus creating a deadly cycle. The use of wire-mesh traps has significantly increased this problem, since these traps last longer before eventually corroding.
Photo by Tomas and Anmarie Tomascik.
The unrivaled splendor and wealth of forms and the delicate tints of the coral structures, the brilliant colours of fishes, clams, sea anemones, worms, crabs, star fishes and the whole rest of the reef animals are so attractive and interesting that it seems impossible to give an adequate description of such a profusion of serene and fascinating beauty. —UMBGROVE 1939A
The above quotation immediately conjures up an image of a beautiful "Pacific atoll" dotted with numerous coral cays, and white sandy beaches lined with tall palm trees swaying gently in the tropical breeze. The year was 1928, and Umbgrove was describing a thriving coral reef community surrounding a small "resort island" called Nyamuk Besar (formerly Leiden), located 6.6 km from Jakarta (Batavia), in Jakarta Bay. By 1985, the live coral coverage of this once thriving coral reef community was given as 1% (Scoffin 1986; Moll and Suharsono 1986), and the various colourful organisms described by Umbgrove were mostly absent. Nyamuk Besar is no longer an island resort. Recent studies of the coral reefs in Jakarta Bay have shown that the once flourishing and diverse coral reef communities (Verwey 1931a; Umbgrove 1939a) are now basically 'functionally' dead systems, undergoing rapid structural degradation mainly through bioerosion and sedimentation (Harger, 1992; Tomascik et al. 1993; Suharsono and Tuti 1994). The animal-algal symbiont associations, characteristic of coral reef ecosystems, have been replaced by a benthic heterotrophic community dominated by filter- and deposit-feeding benthos, characteristic of the muddy environments that surround the island chain (Tomascik et al. 1993).
Wilson (1992) described coral reefs as "...marine equivalents of rain forests…" and "…fortresses of biological diversity in the shallow tropical seas…" It is, there fore, not surprising that the study of these underwater counterparts of tropical rain forests has attracted considerable attention and sentiment. In fact, the future survival of coral reefs has become a worldwide management and conservation issue. Without a doubt, the main threat to the future survival of many coral reef ecosystems, especially the reefs in close proximity to population centres, is pollution associated with rapid economic development and the overexploitation of reef resources by the ever-expanding coastal populations.
The worldwide decline in the ecological state of coral reefs and associated ecosystems (i.e., seagrass beds and mangrove forests), has become a serious concern to managers, scientists, government officials and the general public (Crosby et al. 1995). The worldwide degradation of coral reefs is continuous and systematic, spreading like a plague from human population centres. Since 1991, there were three major international scientific symposia, where the rapid degradation of coral reef ecosystems, throughout much of their distributional range, was the major topic of discussion. Perhaps the two most important, in which Indonesia received considerable attention, were the 1992 Seventh International Coral Reef Symposium, held in Guam; and the 1993 meeting of coral reef experts on "Global Aspects of Coral Reefs: Health, Hazards and History" held at the University of Miami. The 1993 Miami meeting was perhaps the most influential in focusing the attention of the world body to the serious environmental problems that threaten the survival of many coral reef ecosystems and associated habitats. Indeed, in 1991 the Indonesian coral reefs were classified as critical, meaning that if the current trends in overexploitation and mismanagement continue unchecked, the majority of coral reefs in Indonesia (including the eastern regions) will be lost in 10 to 20 years (Wilkinson 1993). The same prognosis was reiterated during the 1993 meeting in Miami (Wilkinson et al. 1994), and again in a 1994 IUCN/UNEP/IOC publication, entitled Reefs at Risk (IUCN 1993). The serious nature of coral reef degradation in the Florida Keys, and the need for immediate action, prompted the United States government to initiate the development and implementation of an inter-agency program called the "Coral Reef Initiative" (CRI). The objective of the U.S.CRI is to coordinate domestic and international efforts aimed at the effective conservation and management of coral reefs and associated ecosystems, seagrass beds and mangroves (Grigg 1994; Crosby et al. 1995). The realization that environmental integrity and ecological stability of coral reefs and associated ecosystems are essential to the well-being of coastal communities, as well as for the maintenance of biodiversity of these systems, prompted other governments to join the U.S.CRI initiative, and to initiate the development of an international coral reef program. The "International Coral Reef Initiative" is supported by the governments of the U.S., Japan, Australia, Jamaica, France, the United Kingdom, and the Philippines. The Indonesian Coral Reef Initiative, through the "Coral Reef Rehabilitation and Management Project" (COREMAP), is a key addition to the International CRI, since Indonesian coral reefs may account for 10% to 20% of the world's total coral reef area. The maintenance of environmental and ecological integrity of Indonesian coral reefs is of global significance, since it is likely that the coral reefs of the Indonesian Archipelago act as long-term net sources of species diversity for peripheral regions (Veron 1995). However, it is also important to realize that at certain times, depending on the general surface circulation patterns in the region, the archipelago may act as a sink, thus receiving significant diversity contributions from the peripheral regions (e.g., Great Barrier Reef, Philippines, etc.). The alternate shifts, from source to sink, may be a crucial factor that may contribute to the high diversity of the Indonesian Archipelago.
Some knowledge of the physico-chemical and biological conditions, under which coral reefs develop and flourish in the various regions of the archipelago, is essential for the development and implementation of appropriate coral reef management and conservation programs. There is an alarming lack of quantitative data on the ambient physico-chemical characteristics and the regional distribution of the various coral reef ecosystems in the archipelago. Until an environmental and biological coral reef database is available, a conservative approach to coral reef management (i.e., people management) is essential. Since coral reef communities in the archipelagic seas have developed in regions where the environment is relatively stable (i.e., small fluctuations in temperature and salinity, and low-intensity disturbances such as storms or earthquakes), it is more than likely that they may lack the functional or structural plasticity (i.e., robustness) to tolerate, at the molecular, organismal or population level, or to adapt to the physical and/or chemical stresses generated by various anthropogenic activities (e.g., urban, industrial and agricultural wastes; oil and mining; forestry; shipping; fishing; tourism; etc.).
Coral reef management and conservation issues in Indonesia are complex, and vary from region to region. However, it is becoming apparent that coral reef education and awareness programs alone will not be sufficient to stem the tide of reef degradation. Innovative management techniques are needed that will focus on people management at the village level, rather than purely on coral reef management. Can we manage a reef? Perhaps, but we know it is possible to manage people and their actions. The issues that need immediate attention are overpopulation and the overexploitation of reef resources by a rapidly growing population without an alternative means of support. Providing new alternatives (e.g., marine polyculture of algae, invertebrates and fish; handicraft; ecotourism; more efficient use of land-based resources, etc.) to traditional exploitative practices must be one of the key components in all coral reef management and conservation programs. One of the key requirements in providing alternatives is that the interventions must be economically viable. For example, a small food industry (nutmeg jam and soft drinks) in the Banda Islands was recently created to develop new products from the nutmeg, thus rejuvenating the ailing nutmeg plantations which have traditionally supported the local economy.
DISTRIBUTION OF CORAL REEFS
World Distribution Patterns
The entire absence of coral reefs in certain large areas within the tropical seas, is a remarkable fact.—DARWIN 1842
Darwin (1842) was the first to publish a detailed map of the worldwide distribution of coral reefs. The map illustrated that coral reef development was restricted to tropical seas, roughly between the 30° N and 30° S latitudes. The most striking feature of Darwin's map was the noticeable absence of major coral reef development in the tropical regions along the western margins of the American and Australian continents, as well as a peculiar absence of barrier reefs and atolls from the Indonesian Archipelago (fig. 4.3). However, it is now well established that extensive coral reefs do occur along the west coast of subtropical and tropical Australia (e.g., c. 100 km2 of reef at Houtman Abrolhos, 29° S), due primarily to the modifying influence of the warm Leeuwin Current, an unusual poleward-flowing eastern boundary current which inhibits upwelling (Hatcher 1991, 1994). The Leeuwin Current originates in the Timor Sea, and is the continuation of the Indonesian Throughflow. Even though the reefs along the west coast of Australia were not marked by Darwin, he recognized that coral reefs were present in the region: "…there are many extensive coral-reefs skirting, often at considerable distances, the N. W. shores…". The confirmation of the existence of barrier reefs and atolls in the Indonesian Archipelago came much later.
The explanation for the general distribution pattern of coral reefs, so clearly illustrated in Darwin's map, had, however, eluded Darwin in his 1842 publication. Even though Darwin (1842) commented on the warm Gulf Stream as a possible explanation for coral reef development in Bermuda (32° N), he did not pursue the question on the lack of coral reefs along the western margins of continents further. It seems that Darwin was thrown off the temperature-control hypothesis by insufficient oceanographic information.
I at first attributed this absence of reefs on the coasts of Peru and the Galapagos Islands, to the coldness of the currents from the south, but the Gulf of Panama is one of the hottest pelagic districts in die world.—DARWIN 1842
Smayda (1966) has demonstrated that high primary productivity in the Gulf of Panama is largely attributed to a seasonal upwelling generated by strong offshore winds as well as by shallowing of the thermocline in the shelf waters. Had Darwin known, as we do today, that the Gulf of Panama is influenced by seasonal upwelling events, which significantly reduce surface seawater temperatures (Glynn and Stewart 1973), the absence of well-developed coral reefs in the Gulf would have been less of a puzzle. Furthermore, the El Niño Southern Oscillation phenomenon, that greatly influences the structure of coral reef communities along the Pacific coast of Panama and the Galapagos Islands, was not known at the time.
Based largely on Darwin's distribution map, Dana (1843) appeared to have been the first to consider temperature as one of the key factors controlling the worldwide distribution of coral reefs. As coral reef research progressed, it was realized that coral reef systems can develop outside the general latitudinal boundaries, provided that they fall within the approximate 18°C isotherm of average winter temperature (Vaughan 1919; Wells 1957; Veron 1986). Recent coral reef distribution maps (not to be confused with generic or species distribution maps) differ little from Darwin's original map. The main difference between Darwin's original map and our current knowledge of coral reef distribution patterns is related to the latitudinal limits of coral reef development, which also includes marginal reef development along the western coastlines of Central America. It is now well established that vigorous reef development or reef accretion, with few exceptions, occurs only in the tropical and subtropical regions of the Atlantic, Indian and Pacific Oceans, roughly between the Tropics of Cancer (latitude 23.5° N) and Capricorn (latitude 23.5° S).
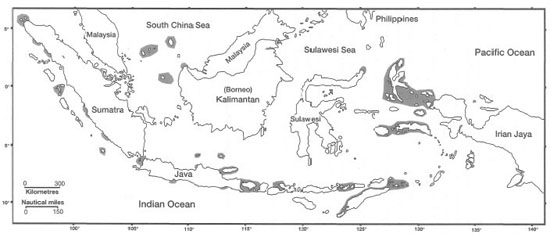
Figure 4.3. Distribution of coral reefs in the Indonesian Archipelago after Darwin (1842).
Source: Darwin 1842.
In the discussion of worldwide distribution patterns of coral reef ecosystems, it is instructive to distinguish between the geographic distribution patterns of scleractinian coral communities and the biogenic structures we call coral reefs, in which zooxanthellate corals (i.e., scleractinians with dinoflagellate endosymbionts) are the dominant, but not the only, constructional component. This is an important point, since reef-building coral communities have a wider global distribution pattern than the carbonate structures which they build (Veron 1986). In the Indo-Pacific some reef-building coral communities occur as far south as the shores of southeast Australia (Veron 1986), and as far north as the east coast of Honshu, Japan (Veron 1992).
These high-latitude locations lie well to the south and north of regions where active coral reef development is presently occurring. Thus, on a global scale we find that a few reef-building coral communities may survive at high latitudes, but do not form reefs due to marginal conditions (Darwin 1842; Veron 1986). Whether these high-latitude coral assemblages are the remnants of former flourishing coral reef communities, or opportunists seeking to colonize new areas, is not fully understood. However, the presence of extensive raised fossil reefs outside of Tokyo suggests that climatic conditions in the past may have been more favourable to reef development in the area (Veron 1995). The distinction between coral reefs and coral reef communities is also applicable on regional (>1000 km2) and local (<1000 km2) scales, especially in Indonesia, where highly diverse zooxanthellate coral communities exist as small outcrops in marginal or unstable environments not favourable for extensive reef development through the Holocene (van Woesik 1994). In Indonesia, however, the distribution of scleractinian corals and coral reefs is related primarily to differences in salinity, turbidity (i.e., light) and sedimentation.
Coral Reefs of the Atlantic Ocean
The Indo-Pacific and Caribbean biogeographic provinces are the two main centres of coral diversity and vigorous coral reef development. While the Caribbean Province is much smaller in area than the Indo-Pacific, it has a similar regional concentration of reefs to the Indo-Pacific. However, major differences between Indo-Pacific and Caribbean biogeographical provinces occur in terms of species diversity, and, to a lesser extent, diversity of reef types. On both accounts, the Caribbean region is less diverse than the Indo-Pacific (e.g., c. 65 scleractinian species vs. 450, respectively). Whether true atolls (i.e., in the Darwinian sense) occur in the Caribbean has often been questioned by geologists; nevertheless, there are at least 10 "atolls" as well as a number of barrier reefs (Milliman 1973). In the southwest Caribbean, coral reefs extend from Colombia along the Central American shelf margin, towards the Yucatan Peninsula, where barrier reefs have developed along the coastlines of Nicaragua, Honduras and Belize. The most extensive barrier reef complex in the Caribbean is located off the coast of Belize along the eastern shelf of the Yucatan Peninsula (Adey 1977). The 250-km-long barrier reef complex runs parallel to the coastline along a south-north axis, with a lagoon that is 10-40 km wide (Rutzler and Macintyre 1982). The "Belize Barrier Reef has often been cited in popular literature as the second-largest barrier reef in the world (Gray 1993). In fact, the largest, or longest, continuous barrier reef runs along the southwest coast of New Caledonia in the western Pacific Ocean, and is about 800 km in length. New Caledonia also has a second barrier reef that parallels the northeast coast of the island. The total length of the barrier reef complex in New Caledonia (i.e., Grande Terre) is about 1600 km (Porcher 1993).
Caribbean atolls are also concentrated in the western region. The best-developed atolls are located just a few kilometres to the east of the Belize Barrier Reef, notably the Turneffe, Lighthouse and Glovers Atolls (Stoddart 1962). The Roncador and Serrana Atolls are located off the coast of Nicaragua. Most active reef development in the Gulf of Mexico occurs in the southern regions on the Campeche Bank, along the western coast of the Yucatan Peninsula. In the eastern Caribbean, the high islands of the Greater and Lesser Antilles are fringed with well-developed fringing reefs, which are the dominant reef formations in the Caribbean. The Brazilian Abrolhos, off the east coast of Brazil (18°30' S) delineate the southern limit of coral reef development in the Atlantic, while the Bermuda Platform, at a latitude of 32° N, delineates the northern limit. The eastern margin of the tropical Atlantic Ocean, along the western coastline of Africa, is devoid of true coral reef development; however, depauperate coral communities can be found at the Cape Verde Islands and in the Gulf of Guinea (Laborel 1974).
Coral Reefs of the Indian Ocean
The Arabian region, including the Red Sea, forms the northwest boundary of Indo-west Pacific reef development. Sheppard et al. (1992) have given an excellent review of the ecology of the marine environment in the Arabian region. The northern limit for coral development is in the northern Gulf of Suez. The dominant reef type throughout the gulf, as in the Indonesian Archipelago, is the fringing reef. In some regions, fringing reefs extend uninterrupted for tens to hundreds of kilometres along the coastline; however, the long-held belief that the world's longest uninterrupted fringing reef (i.e., 4500 km long) is located along the Red Sea coastline (Ladd 1977; Mann 1982; Longhurst and Pauly 1987) has been recently refuted (Sheppard et al. 1992). The Arabian region is tectonically active and, therefore, shares some interesting similarities with the Indonesian Archipelago. The tectonic nature of the Arabian region has been particularly troublesome in determining whether certain reef formations are true atolls or not. Semantics aside, it seems that both atolls and barrier reefs do occur here (Guilcher 1988; Sheppard et al. 1992).
Further to the east, the vast tropical regions of the Indian Ocean are largely devoid of coral reefs. The absence of coral reefs from the central Indian Ocean is related to the absence of suitable substrates, which is a function of bottom topography and geologic history, as well as due to cold upwelling. In areas where submarine ridge systems are present, coral reefs are well established either as fringing reefs, barrier reefs or atolls. In general, the geomorphology and ecological complexity of coral reefs in the Indian Ocean is comparable to the Pacific (Stoddart 1973), as is their generic diversity (Veron 1995). Extensive fringing reefs have developed along the eastern coast of Africa (i.e., from Somalia to the northeast coast of South Africa) as well as around Madagascar; however, they are absent in places because of river discharge. High oceanic islands, such as the predominantly granitic Seychelles and the volcanic Comoros and Mascarene Islands, have well-developed fringing reef complexes (Stoddart 1973). Fringing reefs are well established around the Andaman and Nicobar Islands, in the southeastern Bay of Bengal. However, there are only a few minor barrier reefs present in the Indian Ocean. One of the most studied, and best developed, is a 150-km-long barrier reef that surrounds Mayotte Island in the Comoros (fig. 4.4) (Guilcher 1988; Porcher 1993).
In a recent publication, Wells et al. (1995) mention a 360-km-long barrier reef running along the west coast of the Andaman and Nicobar Archipelagoes, which separate the Andaman Sea from the Bay of Bengal. Perhaps the best known coral reefs in the Indian Ocean are the Maldives, which comprise the highest concentration of atolls in the Indian Ocean. The Indian Ocean also contains a number of extensive submarine carbonate platforms that are entirely below the sea surface, the Great Chagos Bank (13,500 km2) is a good example (Stoddart 1973). Sheppard (1980) reported that on Peros Banhos and Salomon Atolls located on the Chagos Bank, cover of hard substrate by coelenterates averages 80% between 3 and 42 m, and calcareous algae dominate above (e.g., Porolithori) and below these depths. In terms of calcification, corals lose their dominant position to calcareous algae outside this depth range (Sheppard 1980).
Coral Reefs of the Pacific Ocean
The central and western regions of the vast Pacific Ocean are dominated by deep- oceanic reefs and high volcanic islands. The dominant carbonate platforms in these regions are the "Darwinian" atolls, which seem to abound especially in the eastern region of the central Pacific. The highest regional concentration of atolls (76) is located in the Tuamotu Archipelago, which caps the Tuamotu Ridge (Guilcher 1988). As mentioned earlier, the world's second largest barrier reef is in New Caledonia, in the western Pacific. Bora Bora in French Polynesia can serve as a classic example of the "Darwinian" barrier reef (fig. 4.5). Comparable reef types in Indonesia are Pulau Taupan in the Gulf of Tomini, and Pulau Parang off the southeast coast of Seram.
In the Pacific Ocean, true coral reef development occurs to about 29° N; how ever, hermatypic coral communities exist as far north as Tateyama (outside Tokyo Bay), Japan (35° N) (Veron 1992). The presence of these coral communities this far north, and the extensive reef development to the south, is attributed to the direct modifying influence of the Kuroshio, a warm western boundary ocean current. The source water of the Kuroshio is the warm western Pacific pool water located to the east of the Philippine and Indonesian Archipelagoes. Sea surface temperatures of this immense warm pool of tropical water average 28°C.
The southern limits of coral reef development extend as far south as the Lord Howe Islands (31° S) in the Pacific, and to the Houtman Abrolhos (29° S) off the coast of Western Australia, which represent the southern limit of reef development in the Indian Ocean. Lord Howe Island is affected by the warm East Australia Current that is a continuation of the South Equatorial Current, while the Houtman Abrolhos are influenced by the warm Leeuwin Current (Crossland 1988; Cresswell 1991), which is an extension of the Indonesian Throughflow.
In contrast, the absence of coral reefs along the western tropical coastlines of Africa and South America is attributed to strong upwelling that brings cold water masses to the surface, and to the influence of cold ocean currents originating in the Southern Ocean. For example, the cold Benguela Current along the southwest coast of Africa and the Humboldt Current along the west coast of South America are considered to be the main impediment to extensive reef growth. Thus, while the Galapagos Islands are located on the equator in the eastern Pacific, the relatively cool sea surface temperatures associated with the Humboldt Current, and strong regional upwelling, effectively retard extensive coral reef development, even though a number of hermatypic corals are found there. Recent mass mortalities of Galapagos coral reef fauna, a result of the unusually strong 1982-1983 ENSO event, illustrate that eastern Pacific coral communities, adapted to cooler environmental conditions, are greatly influenced by large-scale atmospheric-oceanic perturbations, which periodically increase sea surface temperatures 2-3°C above normal.
Figure 4.4. French Mayotte Island in the southeastern Indian Ocean (northwest of Madagascar), Comoro island chain. The barrier reef is most extensive along the east and northeast coast of the volcanic island. The dark shade signifies vegetation. Light sections of the barrier reef represent extensive reef flats. The seaward edge of the barrier reef complex is delineated by white breakers. A coral cay is clearly visible at the southern region of the barrier reef. Image from the SPOT 2 spacecraft (20 m resolution multispectral image enhanced with 10 m resolution panchromatic data).
Courtesy of SPOT Image Corporation. Made from colour print.
Figure 4.5. Bora Bora Island in the central South Pacific Ocean, Society Islands, French Polynesia. A classical "Darwinian" barrier reef surrounding ancient volcanic islands. The seaward edge of the barrier reef complex is delineated by white breakers. The reef rim is between 1-2 km in width. The northern and eastern part of the barrier reef consists of an elevated (0.5-0.7 m above sea level) ancient reef (about 2250 years old) bearing numerous wooded islets (motu) separated by shallow channels (hoa), through which reefal sediments enter the lagoon. The lagoon has only one deep pass (ava), which is 20 m deep (Chevalier 1973). The Teavanui (Great Passage) is located to the west, on the leeward side of the island (Chevalier 1973; Guilcher 1988). Note the extensive sediment fans on the northwest section of the barrier reef, which illustrate the lagoonward transport of reefal sediments. Image from the SPOT 2 spacecraft oh 7 July 1991 (20 m resolution multispectral image enhanced with 10 m resolution panchromatic data).
Courtesy of SPOT Image Corporation.
It is important to stress that other environmental factors, such as light, salinity, turbidity, current patterns and hydrology play an important role in limiting coral reef development on regional and local scales (van Woesik 1994). Since Darwin's monumental work on The Structure and Distribution of Coral Reefs, we have learned a great deal about the mechanisms through which environmental factors, such as sea temperature, salinity, light intensity and day length, exert their influence in determining the global distribution pattern of coral reefs. However, our understanding of coral reef distribution patterns on regional and/or local scales lags far behind.
REGIONAL PERSPECTIVE
Coral Reefs of the Indo-Pacific Region
From the global perspective, most of the world's coral reefs (c. 85%) are located in the Indo-Pacific (Smith 1978). The Indo-Pacific region extends from the shores of east Africa and Madagascar, to the Arabian Gulf and Red Sea, and then east to French Polynesia in the central Pacific Ocean. The widest latitudinal extent of coral reefs in the Indo-Pacific occurs roughly between southern Japan and southern Australia. Veron (1995), using an extensive coral database, has divided the Indo-Pacific coral fauna into six distinct regions based on the geographic ranges of scleractinian genera (i.e., Indo-west Pacific, central Pacific, South-east Pacific, Far southeast Pacific, Far east Pacific and the Hawaiian Islands). The Indo-west Pacific is the largest and most diverse province with a relatively homogeneous distribution of genera extending from the Fiji Islands to the Red Sea (Veron 1995). The Indonesian Archipelago is at the centre of this most diverse province.
However, when we look at the actual concentrations of coral reefs (i.e., number of reefs or the actual area covered), a different picture emerges. According to Potts (1983), the majority (53%) of the coral reefs within the Indo-Pacific region can be classified as continental shelf reef systems, located along the continental margins of Southeast Asia and Australia. Only about 19% of the Indo-Pacific reefs fall within the stereotypical "mid-oceanic reefs" category (e.g., atolls), characteristic of the oligotrophic central and western Pacific Ocean.
The Indian Ocean contains 20% of the Indo-Pacific reefs, which are a mix of marginal shelf reefs as well as mid-oceanic reefs (Potts 1983). As a direct result of geologic history, there is a definite longitudinal gradient in coral reef distribution within the Indo-Pacific region. Within the Indo-west Pacific Province the most extensive reef development, as well as the highest coral species diversity, occur in a vast and geologically diverse area extending from the Andaman Islands in the Bay of Bengal to the Great Barrier Reef along the east coast of Australia, and roughly between 24° N - 24° S (reef development at higher latitudes is here considered as marginal).
THE NATURE OF CORAL REEFS
Recent coral reefs are massive, wave-resistant, biogenic carbonate structures with positive topographic features that were built over millennia, mainly through the action of scleractinian (Scleractinia) reef-building corals and crustose coralline algae. Coral reefs occur in a great variety of forms and sizes, since they are a product of a complex history of interactions between the biotic (e.g., calcification, cementation, consolidation, dissolution and bioerosion) and abiotic (e.g., tectonic setting, sea-level fluctuations, wind, waves, currents, and tides) environmental factors (fig. 4.6).
The concept of what a coral reef is, in physical and ecological terms, is discussed in greater detail by J. van der Land (see box 4.1), while the objective of this discussion is to present basic information on the biological nature of coral reefs and their distribution. The terminology for coral reef classification pre-dates that of Darwin (1842), who formally classified and described the various types of coral reef formations under three broad categories, namely the fringing reefs, barrier reefs and atolls. Darwin's broad reef classification system, based mainly on geomorphological features, withstood the test of time, even though the term coral reef, as the appropriate generic name of Recent (Holocene) carbonate platforms in the tropical seas, has been recently questioned (Hillis-Colinvaux 1986). The terminology challenge centres on which reef-building group (i.e., corals or coralline algae) is the dominant builder of Recent reef structures. It is interesting to point out, however, that it was realized early in the history of reef science that scleractinian corals were not necessarily the main components of modern "coral reefs" (Molengraaff 1929).
An important percentage of many so-called coral reefs is not built up by corals but by other organisms, among which foraminifera and calcareous algae far pre dominate.—MOLENGRAAFF 1929
Since the review of Umbgrove (1947), no serious attempt has been made at a comparative systematic study on the general geomorphology of Recent (i.e., Holocene) Indonesian reef systems. Hopley's (1982) Great Barrier Reef treatise stated that:
Within each class of reef, fringing reef, barrier reef, or atoll, there is much morphological diversity. This is particularly true of the reefs of continental shelves that fall generally within the barrier reef division of Darwin, and nowhere is the diversity as great as in die Great Barrier Reef of Australia.
Based on the meager data available, it seems that the geomorphological diversity of Indonesian shelf reefs alone is comparable to, if not greater than, that of the Great Barrier Reef. The classification system of Holocene reefs proposed by Hopley (1982) is applicable to Indonesian reefs. The six types of shelf reefs, ranging from the unmodified antecedent-platform reefs to senile planar reefs (Hopley 1982) have been identified throughout the archipelago.

Figure 4.6. Fringing reefs are the most familiar and abundant coral reef type throughout the Indonesian Archipelago. These biogenic structures protect thousands of kilometres of coastline, and are a significant, but rarely appreciated, economic asset. Aerial view of a fringing reef along the northwest coast of Bonarate Island, Flores Sea. The fringing reef surrounding the island is 45 km long, with an estimated surface reef area in excess of 25 km2.
Photo by Tomas and Anmarie Tomascik.
Throughout Indonesia, thousands of kilometres of volcanic coastlines are fringed by diverse coral- and coral-associated communities, even though they have not altered the original basement to the point where geologists would consider them as reefs. Nonetheless, from a biological and ecological perspective, these non-reefal communities exhibit all the functional properties of a "classical" coral reef ecosystem. McManus (1988) has made a clear case for distinguishing between reefal and non-reefal coral communities (i.e., carbonate and non-carbonate), since, as he points out, this distinction may have significant implications for the sustainable management of coral-reef resources. There is substantial support for this view, since earlier studies (McManus and Wenno 1981) and the results of the Snellius-II Expedition have clearly demonstrated that in many parts of eastern Indonesia mature and highly diverse coral communities do indeed occupy non-reefal substrates (Best et al. 1985; Best and Boekschoten 1988; Hoeksema and Moka 1989; van der Land 1989). It should'be noted, however, that many non-reefal coral communities may be ephemeral features of many shallow-water coastal environments, where they are strong indicators of environmental stress (van Woesik 1992).
Recent studies on new lava flows suggest that coral reefs, as such, may never develop, or are very slow in developing, in areas where substrate instability (e.g., submarine rock slides) periodically destroys the initial carbonate veneer, and sets the reef growth back to its original phase (Tomascik et al. 1996). Nevertheless, these young scleractinian coral communities, while being geologically distinct from a typical fringing reef, may nonetheless have a similar ecological function as a mature coral community established on structural reefs (i.e., carbonate substrates), and may indeed support the same species, but not the same size classes.
In fact, many non-reefal coral communities seem to support higher reef fish biomass than adjacent reefal communities. The suggestion that diverse coral communities on non-carbonate substrates (i.e., non-reefal communities) are ecologically distinct from coral communities on carbonate substrates (i.e., reefal coral communities) (McManus 1988) has so far not been substantiated by quantitative studies, even though it is intuitively appealing. While there are obvious grounds to view reefal and non-reefal coral communities as distinct ecological entities in scientific investigations, it is suggested that from management and conservation perspectives it may be prudent to treat all productive coral (scleractinian and non-scleractinian) communities as similar entities, so that these productive areas are not excluded from management programs. Too frequently the non-reefal communities are not included in resource survey inventories, which tend to focus only on areas where coral reefs are clearly visible. As a result, potential fisheries resources of the area may be seriously underestimated.
The distinction between coral reefs and coral communities is, however, useful when discussing general (i.e., global and regional) distribution patterns of coral reefs and biogeographical distribution patterns of coral (i.e., scleractinian) communities. It has been clearly demonstrated that the present-day distribution patterns of coral reefs and coral (scleractinian) communities in the central Indo-west Pacific Province and elsewhere are not exactly the same (Sheppard et al. 1992; van Woesik 1992; Veron 1992; Veron and Minchin 1992).
Coral Reef Definitions
One of the main problems in comparing the available coral reef area estimates is directly related to the lack of a precise definition for the term coral reef. As was clearly pointed out by van der Land (1989) and Crossland et al. (1991), the term coral reef has seldom been explicitly defined in scientific studies or in large-scale coastal resource inventories. Note, however, that the geologists have a good number of stratigraphic definitions in place. The need for a working definition of coral reefs was raised earlier by McManus (1988), who stated that the lack of a clear definition has caused a great deal of confusion on how to assess and manage coral reef resources. This is an important management and conservation issue, since many high-diversity coral reef communities (i.e., corals and the associated fauna and flora) in the Indonesian Archipelago flourish on other substrates (e.g., coarse alluvial sediments, volcanic lavas, etc.).
For example, the 1949 eruption of Ruang Volcano, Sangihe Archipelago (North Sulawesi), was accompanied by a large lava flow that originated from the main crater (fig. 4.7). The lava flow destroyed a considerable section of a fringing reef to a depth of about 60 m. A brief visit to the lava flow in 1993 has provided valuable new insight on coral colonization of andesitic lava flows in areas with high local coral species diversity and environmental stability, excluding volcanic activity. The 1949 lava flow can be considered as an ecologically functional fringing reef, in terms of coral abundance, coral cover and coral species diversity. In addition, the diverse coral community (between 125-175 scleractinian species; rapid assessment) supports a full complement of associated reef flora and fauna (Tomascik and Mah, pers. obs.) (fig. 4.8). Unfortunately, the thickness of the carbonate deposition since the eruption could not be ascertained.

Figure 4.7. Andesitic lava from the 1949 eruption of Ruang Volcano, Sangihe Archipelago, North Sulawesi.
Photo by Tomas and Anmarie Tomascik.
After 46 years of colonization and succession of communities, the Ruang lava flow - while being an "embryonic" reef from a geological perspective (i.e., accumulation of CaCO3) - is clearly a mature coral reef community (high abundance of massive Pontes spp. up to 1 m in diameter) from an ecological perspective.
Satellite remote sensing technology, introduced in the early 1970s, is only now making major inroads into coral reef science and coral reef management (see fig. 4.5). It is, therefore, imperative that a working definition of the term coral reef he adopted, so that data collected in the future throughout the Indonesian Archipelago, and other regions of the world, can be properly analysed, meaningfully compared and used to develop and implement sound management and conservation policies.
Box 4.1. What is a coral reef?
J. van der Land, National Museum of Natural History, Leiden, the Netherlands.
To most people, a coral reef is an aesthetically appealing assemblage of sedentary animals and plants surrounded by colourful fish in crystal-clear tropical waters. Therefore scientists, conservationists and managers tend to use the term coral reef in a similar sense in non-scientific contexts, especially for public relations and fund-raising purposes. I do not oppose this and if I did it would be in vain. However, this loosely and subjectively defined term is being used in scientific texts as well, often with the non-scientific meaning but also with a number of other meanings and connotations. This has led to considerable confusion in the science, management and conservation-oriented activities.
The English word i'reef originated from the Dutch rif, a term used by the old seafarers to indicate an elongated, rocky shallow, arising from deep water. To them a reef was a shipping hazard. In science reef has many different meanings and the term coral reef is particularly confusing. In the earth sciences a coral reef is a limestone structure partly consisting of coral skeletons. Up until the present day there still is a general state of ignorance about reef genesis and development. As a result most earth scientists seem to prefer neutral terms such as carbonate platform. Paleontologists found that reef limestone may originate from only a small percentage of stony corals or even from completely different organisms. In addition, coral assemblages often occur in non-reefal situations.
In marine biology the term coral reef has a variety of meanings. Sometimes it refers to a carbonate platform or a part of it, provided living corals are present, but it is also quite often used to indicate a benthic community in which stony corals dominate. In most cases the term is used loosely and it may be difficult to determine which definition is used by an author. A complicating factor is that Caribbean reef environments differ in several respects from Indo-Pacific reefs. Some biologists avoid the term coral reef replacing it with coastal hard-substratum communities or tropical marine coastal ecosystems. Botanists have generally disliked the name coral reef and even proposed the term algal reef. In fisheries biology much confusion has arisen from the unspecified use of the coral reef concept. In my opinion, marine biologists should avoid confusion, at least in scientific texts, by not attaching any biological meaning to the name (coral) reef and by using it in a geomorphological sense only (i.e., to indicate a carbonate platform).
Reef Communities
Quite often the term coral reef is used, even by marine biologists, to indicate a certain benthic community or ecosystem which equates fairly well with the layman's perception. Others mean the same when they use terms such as THE coral reef ecosystem. However, even if we restrict the meaning to scleractinian-dominated communities there is no such thing as THE coral reef: there are several totally different types. Abstractions are needed in science, but when they are oversimplifications they are counterproductive. A disadvantage of such a simplification is that reef ecologists tend to consider the results of their studies to be generally applicable.
Above I used the term community (biocoenosis) and I continue to do so throughout this paper because of its heuristic value. Several authors tend to use terms like association or assemblage instead of community, probably because they doubt the reality of communities. The word assemblage already has several Box 4.1. (Continued.) different meanings. There is nothing against the use of community or biocoenosis as long as their restricted value is realized. In this respect we tend to be a little careless. A biocoenosis does not exist in nature but is a mental sample of organisms at a given time at a given place (only the gradients of exogenous factors causing the distribution of the species are real). Similarly an ecosystem is a mental sample of the biota and the totality of interactions and exogenous factors. If the sample is too large, as in the example mentioned in the first paragraph, it does not make sense.
I prefer a broad definition of the term (coral) reef. In this sense, a reef, and particularly a complicated one like an atoll, may harbour a variety of hard- and soft-bottom communities. On hard-bottoms there are several types of scleractinian-dominated communities, but also communities in which other groups, such as calcareous algae, alcyonarians and sponges, dominate and also vegetations of macro-algae or filamentous epilithic algae (algal turf). Certain soft-bottom communities such as seagrass beds are usually present on Indonesian reefs. Mud flats, often with mangrove stands, are most extensive in areas exposed to considerable terrigenous influences. Apart from benthic biocoenoses there are also demersal and often even terrestrial reef communities. Even professionals tend to link hermatypic corals with reefs, without realizing that scleractinians often occur in non-reefal situations. Some of the most diverse coral-dominated communities in Indonesian waters occur on hard bottoms of volcanic origin and many species abound even in sandy lagoons or on muddy slopes.
Diversity
Coral reef communities are generally thought to be extremely diverse and this is one of the reasons why they are often considered the marine counterparts of tropical rain forests. Among marine communities the highest species diversity undoubtedly occurs in certain reef communities and this is particularly the case in Indonesian waters, which form the centre of diversity of several groups of marine organisms. However, several types of reef communities, even scleractinian-dominated reef-crest communities, are not diverse at all. Monospecific stands of stony corals and communities with very few dominant species are quite common.
Eastern Indonesia may be the centre of marine diversity, but until now this has only been a statistical, zoogeographical conclusion. Several historical, oceanographical, climatological and geographical factors have coincided to make this a species-rich area. The richness in types of communities, benthic and demersal species and symbiotic relationships is probably due to the extremely varied geography of the archipelago, with sheltered as well as exposed conditions and with estuarine as well as oceanic waters. The identification of diversity is of great interest from a conservation point of view. We want to know how diversity can be observed in the field. The human eye first appreciates the aesthetic value of certain communities, but this is not the same as natural richness. Superficial observations are insufficiently accurate to determine the status of benthic biota. In some cases poor-looking benthic communities are very rich in species. Several communities consist of small numbers of species only, but many different communities of this type in a small area can make that area as a whole extremely diverse. Obviously we have to account for within-habitat diversity (alpha diversity) as well as between-habitat diversity (beta diversity).
Stability
Another popular conception is that coral reef communities are very stable, rather like rain forests, with giant coral colonies growing quietly for ages. However, the concept of reef stability is not generally correct and totally undisturbed reef communities may even be an exception. Diversity in reef communities may be retained because of regular disturbances, which is well in accordance with the general intermediate disturbance theory on biological diversity. Acroporids and poritids are the most important scleractinian reef builders, but in totally different ways. Porites and other species with massive colonies may contribute directly to the reef structure, while acroporids do this in an indirect way. Branching and foliose corals, completely dominating in many localities, are easily fragmented but are usually capable of rapid regeneration. For such corals fragmentation may even be the most important means of reproduction. The complete destruction of an /Acropora-stand by a storm, sometimes blamed on fish bombing, is part of a natural chain of events. Usually we lament the complete overgrowth of coral stands by green algae and we tend to consider such an event "abnormal". It is often ascribed to human interference such as overfishing of grazers or human-induced eutrophication, but this phenomenon also occurs in pristine locations. Destruction and subsequent building of ramparts by storms, sometimes even above water, may well be just as essential for the maintenance and growth of a reef as the growth of calcareous frameworks. We know that several types of phenomena, such as El Niño events, cyclones, Acanthaster outbreaks, bleaching, and diseases may be destructive. Entire coral stands may be killed, but it is difficult to determine if the cause was human-induced or a natural phenomenon, which is of significance for the management of benthic biota.
Catastrophic disturbances are rare, but we have to realize that rare phenomena have a relatively great influence because communities do not adapt. Paleontological studies have shown that enormous biological changes may take place over relatively short periods of thousands of years, such as those caused by changes in temperature and sea level. Regrettably man is now capable of causing catastrophic disturbances as well as more gradual global changes over much shorter periods. Tropical benthic communities may be resilient to instabilities and be capable of regeneration to a much larger extent than we are aware of, but anthropogenic pressures are increasing.
Nutrient cycling
It is a generally-held view that coral reefs are oases in the desert of the nutrient-poor tropical oceans. Comparisons with tropical rain forests, because of the high primary production due to retention and recycling of nutrients, are also popular. However, such conceptions describe the situation on oceanic reefs, and consequently they are not generally applicable. Most reefs are not situated in purely oceanic conditions and may be exposed to considerable terrigenous influences. This certainly holds true for the numerous reefs in Southeast Asian seas, situated where more than half of the world's river sediment loads are deposited.
Reef fisheries
Tropical reefs support some of the highest standing crops of fishes, consisting mostly of planktivores, small herbivores and primary carnivores. Harvesting tends to concentrate on high-order carnivores and recovery of depleted stocks may be slow. Therefore signs of overfishing are apparent, emphasising the need for fisheries management.
In fisheries biology the use of the term coral reef is ambiguous. Some authors use the term as I defined it previously, many use it to indicate scleractinian-dominated communities, but most seem to lack a well-defined concept. I have the impression that reef fishery is mostly considered to be the activity of fishermen working in reef areas, but fishery biology studies (populations, life cycles, recruitment) tend to be restricted to demersal fish living in coral-dominated communities. This biased approach gives an inadequate impression of the economic potential of reef fisheries. Reef fisheries can at least partly be a type of pelagic fishery and therefore of economic importance.
In several areas fishermen stated that from their point of view, seasonal pelagic fishing with lines or lights from small boats is the most financially important. This may relate to valuable species of fish such as tuna, as well as the substantial squid fishery. Even a small reef island provides space for a village of up to 1000 inhabitants, or it may serve as a temporary base. The reefs provide safe anchorages as well as food during the bad weather. Coral islands form a complete base for pelagic fisheries. I once compared them with factory ships, waiting for migratory fish schools rather than actively hunting them. I have the impression that there is a potential for further development of "reef fisheries" for pelagic stocks if proper attention is given to socioeconomic aspects such as boat building, post-harvest treatment and marketing procedures.
We may conclude that there is considerable confusion over nomenclature in reef research. There is more than one answer to the question of "What is a coral reef?". Efforts should be made to arrive at some concensus on a concise definition acceptable for general usage and by the various scientific fields.

Figure 4.8. A diverse coral community on a 1949 lava flow supporting high biomass and diversity of all reef-associated organisms. Ruang Island, Sangihe Archipelago, North Sulawesi.
Photo by Tomas and Anmarie Tomascik.
Coral Reef Definitions for Management
In their review of global coral reef productivity, Crossland et al. (1991) used an explicit definition to delineate the outer boundaries of coral reefs. Their definition was based on a number of biogeochemical criteria that are relatively easy to identify; thus they are valuable from both management and conservation perspectives. Since light is an all-important environmental factor controlling reef accretion in the tropical regions, the bottom of the euphotic zone was taken to represent one of the boundary criteria. The depth of the euphotic zone varies greatly throughout the Indonesian Archipelago, mainly along a longitudinal gradient. There is a pronounced light attenuation gradient moving in a westerly direction from the Flores Sea to the western Java Sea as well as from the Banda Sea to the Arafura Sea and the coastal waters of Irian Jaya (Gieskes et al. 1989). For example, the euphotic zone in the Banda Sea may extend below 100 m during the Northwest Monsoon (i.e., non-upwelling period), and is significantly deeper than in the Java Sea, where water turbidity is high year-round. In the Flores and Banda Seas vigorous coral reef growth may extend to depths of 60 m or more, while in the western Java Sea it is restricted to depths of less than 15 m (Verwey 1931a; Tomascik and Mah, pers. obs.). Therefore, the use of a single average value for the euphotic zone to delineate the outer boundary of coral reefs in the various parts of the Indonesian Archipelago may not be appropriate. It may be more appropriate to define the reefal boundaries at regional, or preferably at local levels.
The second criteria used by Crossland et al. (1991) to define coral reef areas was the nature of sediment composition. They proposed that the outer boundary of coral reef areas be delineated by a transition to less than 80% reefal (i.e., carbonate) sediments. This value is appropriate for oceanic and offshore reef systems, but not necessarily for coastal systems influenced by terrestrial processes. For example, the Berau Islands in East Kalimantan are located in close proximity to the Berau River, which releases large quantities of silt year-round. Surprisingly, there are a number of extensive fringing reefs to the south and north of the river delta, as well as a barrier reef and three atolls within a few kilometres of the river. The three atolls (Kakaban, Maratua and Muaras) are sufficiently far offshore (>60 km) that they are situated in mostly oceanic conditions. However, the numerous patch reefs and extensive shallow-water platform reefs (e.g., P. Panjang with a 143 km reef flat) that comprise the barrier reef system are under the direct influence of the Berau River as well as the Bulungan River, 60 km to the north. The sediments of the inshore fringing reefs contain relatively high percentages of fluvial deposits. The reef flats exhibit distinct lagoonal characteristics, supporting high abundances of burrowing macroinvertebrates (fig. 4.9). The reef flats are extremely productive coral reef habitats, and even with high percentages of fluvial deposits, they clearly are of reefal origin and should be classified as such.
On many volcanic islands (e.g., Solor, Adonara, Lembata) the reefal carbonates may constitute less that 50% of the coastal sediments, even though the entire island may be fringed by a reef. In other areas, sediments along large sections of coastline may be mostly of non-reefal origin (e.g., Merdeka Bay, Lembata Island; Luwuk, Central Sulawesi), even though well-developed fringing reefs are present.
Taking these, two criteria (i.e., light and sediments) under consideration (the third criteria stipulating >50% dominance by tropical organisms is not relevant here), Crossland et al. (1991) delineated the outer boundary of coral reefs as: "...the bottom of the euphotic zone or transition to <80% reefal sediments (whichever comes first)…". The definition includes outer reef perimeters, Halimeda bioherms as well as the extensive inter-reefal areas of atolls and barrier reef systems. However, the definition does not include the non-reefal coral reef communities, including scleractinian communities on non-carbonate substrates. In die eastern part of Indonesia, which is tectonically very active, long stretches of non-carbonate (e.g., volcanic) coastlines are covered by luxuriant coral communities with a large complement of coral-reef-associated fauna (fig. 4.10). In fact, many of these non-reefal coral communities support higher fish biomass than comparable reefal habitats, as well as a higher diversity of relatively rare coral species (Best and Boekschoten 1988). Nonetheless, these communities maybe transitory features of stressed environments (van Woesik, pers. comm.).

Figure 4.9. Subtidal reef flats of inshore coral reefs in the Berau Islands are highly productive habitats with distinct lagoonal characteristics, resulting from the relatively sheltered position of the reef complex, and its close proximity to a major river delta. Raburabu patch reef at high spring tide, East Kalimantan.
Photo by Tomas and Anmarie Tomascik.
Considering the high diversity of coral reef types in the Indonesian Archipelago, the outer boundaries of reefal areas for fringing reefs, barrier reefs, patch reefs and atolls should be defined separately. With regards to fringing reefs, their definition for management and conservation needs to be modified to include non-reefal coral communities. It is suggested that all non-reefal substrates which support a living veneer of predominantly (>50%) scleractinian corals and other calcifying organisms is considered equivalent to a fringing reef. For management purposes, the outer boundary of fringing reefs should be set at the 100 m isobath or transition to <30% reefal sediments, whichever comes first. The 100 m depth rep resents the average maximum depth of commercially important groupers (Serranidae) and snappers (Lutjanidae) on the outer seaward reef slopes in many parts of the archipelago. Because many fringing reefs in the archipelago occur along coastlines heavily influenced by terrestrial runoff (e.g., Sumatra, Java, Bali, Kalimantan, Sulawesi, Irian Jaya), the minimum content of reefal sediments in offshore sediments is set to 30%.

Figure 4.10. The shelves of many volcanic islands in the archipelago, such as Siau, Sangihe Archipelago, are covered by diverse shallow-water coral reef communities.
Photo by Tomas and Anmarie Tomascik.
With regards to barrier reefs, the seaward boundary is set at the 100 m isobath or transition to <30% reefal sediments (whichever comes first), as in the case of fringing reefs. The entire barrier reef lagoon is included in the definition of reefal area, since most barrier lagoons support commercially important fisheries (e.g., groupers). The outer boundaries of atolls and patch reefs are the same as for the fringing reefs and barrier reefs. The lagoonal and reef flat areas of atolls and patch reefs are considered as parts of the reefal area.
GLOBAL CORAL REEF AREA ESTIMATE
While we have a good picture of the worldwide distribution pattern of coral reefs, even though the causative mechanisms have still to be classified, there are no accurate estimates with regards to their actual areal extent. The fact that we still do not have a reliable estimate of the area of coral reefs attests to our still-limited knowledge of these diverse ecosystems. Nevertheless, there are a number of recent coral reef area approximations, each giving a very different value. The highest global estimate is given by Achituv and Dubinsky (1990), who suggested that: "At the present, coral reefs cover some 2 x 10 km of tropical oceans…" However, no information is provided on the methodology of the approximation, or on the definition of "coral reefs". The lowest global coral area coverage was given by De Vooys (1979), who used an area of 112 x 103 km2 in his global estimate of coral reef net primary production rates. However, De Vooys' estimate is most likely very conservative.
Smith (1978) suggested that the minimum area of the world's coral reef ecosystems is about 6.17 x 10 km. Of the few global coral reef area estimates available today, Smith's estimate seems to have received the most acceptance (table 4.1) (White 1987; Beatley 1991; Crossland et al. 1991). Other estimates, basically variations of Smith's (1978) model, suggest that coral reefs cover about 0.2% of the world's ocean area (Barnes and Chalker 1990), which would roughly translate into 7.24 x 105 km2 of coral reefs if we take the present-day world's ocean area to be 362.033 x 106 km2 (Menard and Smith 1966).
The latest regional estimate by White (1987), suggests that the fringing reefs of Southeast Asia alone may conservatively comprise about 30% of the world's coral reef area. Since the Indonesian Archipelago has the longest coastline (i.e., 81,000 - 201,000 km) in the central Indo-west Pacific Province, fringing reefs are the dominant coral reef types in the archipelago.
The Great Barrier Reef, stretching for some 2300 km along the continental shelf of eastern Australia, is currently considered to be the largest and most diverse biogenic structure on the planet (Maxwell 1968; Hopley 1981; van Woesik 1994). However, the Great Barrier Reef is not a single continuous reef unit, as the name somewhat misleadingly implies, but rather, it is a series of about 2425 individual coral reefs (Maxwell 1973). In his comprehensive review of the geomorphology of the Great Barrier Reef, Hopley (1982) identified and classified 1301 reefs, and stated that an additional 400 reefs may be found between 9° S and 11° S. Therefore, Hopley's estimate can be viewed as the minimum number, excluding the fringing reefs, whose number, according to Hopley and Partain (1987), stands at 545 with a total area of about 350 km. To put this number in terms of an Indonesian perspective, where fringing reefs are a major coastal feature, consider the fact that the fringing reefs of the Tanimbar Islands alone cover an area of over 900 km2. With regards to barrier reefs, it seems that the barrier reef along the southwest coast of New Caledonia has longer stretches of continuous reef than the Great Barrier Reef, whose outer reefs are at some points (e.g., central region) separated by rather large distances (>50 km). Most of the coral reefs that comprise the Great Barrier Reef are relatively small, usually less than 2.6 km2 in surface area. According to Maxwell (1968,1973) there are only 75 reefs that have an area greater than 50 km2. The actual coral reef area of the Great Barrier Reef was estimated to be between 4300 and 5000 km2 (Maxwell 1973). However, new estimates, based on remote sensing technology (e.g., Landsat, Spot), suggest that the total reef coverage of the Great Barrier Reef is about 20,000 km2 (Hopley and Partain 1987), but Fagerstrom (1988) suggested that the living coral reef covers an area of only 11,700 km , which is roughly 4% of the Great Barrier Reef. These estimate discrepancies may be primarily a result of different coral reef definitions used by the above workers. If considered as a single carbonate province, the Great Barrier Reef covers an area of about 290,000 km2 (Hopley 1982).
Table 4.1. Current estimates of the total area of the world's coral reefs.

AREAL ESTIMATES OF CORAL REEFS IN INDONESIA
The areal extent of coral reefs in the Indonesian Archipelago, as for the rest of the world, cannot be stated with great precision. The current official estimate of coral reef coverage in Indonesia stands at roughly 7500 km2, or 750,000 ha (KLH 1992). It is important to stress, however, that this estimate takes into consideration only the intertidal portion (i.e., exposed at low tides) of fringing reefs, which is usually taken as the distance from shore to the seaward edge of the outer detrital ridge (fig. 4.11). The intertidal reef flat is frequently the area of the coral reef that is depicted in most hydrographic charts with scales 1:1,000,000 to 1:25,000. Obviously, the larger the scale (i.e., the greater the detail), the more accurate are the measurements. The reefal area that is currently given as the official estimate was based on the assumption that about 45% of Indonesian coastline is fringed by coral reefs (i.e., fringing reefs only), which are on average about 200 m in width. While fringing reefs with narrow reef flats and steep seaward drop-offs (i.e., vertical reef slopes) are a characteristic feature of many small and large islands (i.e., Banda Islands, Buru; south coast of Java, Sulawesi, etc.), there are as many islands where the fringing reefs (i.e., reef flats) extend a few kilometres offshore (i.e., Tanimbar Islands, Berau Islands, Salabanka Islands, Aru Islands, etc.). Many continental islands on the Sunda Shelf have wide fringing reefs. For example, the fringing reef that skirts the 15-km-long coastline of Pulau Keramian, northeast Java Sea, extends almost 4 km offshore at the southeast coast of the island (fig. 4.12). In many areas (e.g., East Kalimantan, Tanimbar Islands) the seaward reef slopes of many of these wide fringing reefs have very gentle slopes that can extend the reefal area an additional few kilometres offshore to the 10-30 m isobath, where coral reef communities can still flourish (fig. 4.13).
To illustrate the points being made, consider the following example. From an interpretation of hydrographic charts (scale 1:250,000), we can estimate that Larat Island, just north of Yamdena in the Tanimbar Islands, is fringed by a continuous reef that is about 90 km in length (fig. 4.14). With an average width of about 1 km (considerably more than the 0.2 km national average), the fringing reefs have an area of 90 km. This area represents only the intertidal portion of the fringing reef that becomes exposed during low tides (i.e., reef flat). Almost the entire 410 km coastline of Yamdena Island is fringed by a reef that on the east coast extends virtually uninterrupted for about 170 km. The average width of the exposed reef flat is about 1 km, which translates into about 410 km of reef flat area around Yamdena. However, the reef extends seawards from the edge of the reef flat to the 20 m isobath. The 20 m isobath is on average (based on random sampling of points along the coastline) about 2 km offshore. Thus, using simple arithmetic, the fringing reef surrounding Yamdena has an area of approximately 820 km2. The combined coastline length of the 10 largest islands in the Tanimbars (i.e., Yamdena, Selaru, Seira, Selu, Wuliaru, Wotap, Larat, Maru, Molu and Fordate) is about 900 km. Using the 20-m-depth isobath as the outer boundary of the fringing reef, the approximate area of fringing coral reefs in the Tanimbar Islands is 1800 km2, or 24% of the current national figure.

Figure 4.11. Typical reef flat of a fringing reef. The algal-dominated detrital ridge may extend from hundreds to thousands of metres offshore. This portion of the reef is usually depicted on most hydrographic charts with scales of 1:500,000 or larger. Watubela Island, northeast Banda Sea.
Photo by Tomas and Anmarie Tomascik.
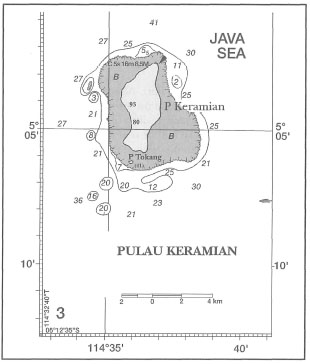
Figure 4.12. Chart of Pulau Keramian in the northeast Java Sea. Depths in metres.

Figure 4.13. Gentle reef slopes are a characteristic feature of many coral reefs in shallow-water areas, such as the Berau Islands. Seaward slope of a 3-km-wide fringing reef off Tanjung Situkan, 24 km north of the Berau River delta, Berau Islands, East Kalimantan.
Photo by Tomas and Anmarie Tomascik.
However, not all estimates are as straightforward as the Tanimbar example. The difficulty in the interpretation of hydrographic charts to obtain coral reef areal estimates can best be illustrated by a simple example from a fringing reef that surrounds Bahuluang (shoulder in the open sea; van der Land and Sukarno 1987), which is a volcanic island off the southwest coast of Selayar, in South Sulawesi (fig. 4.15 A, B).
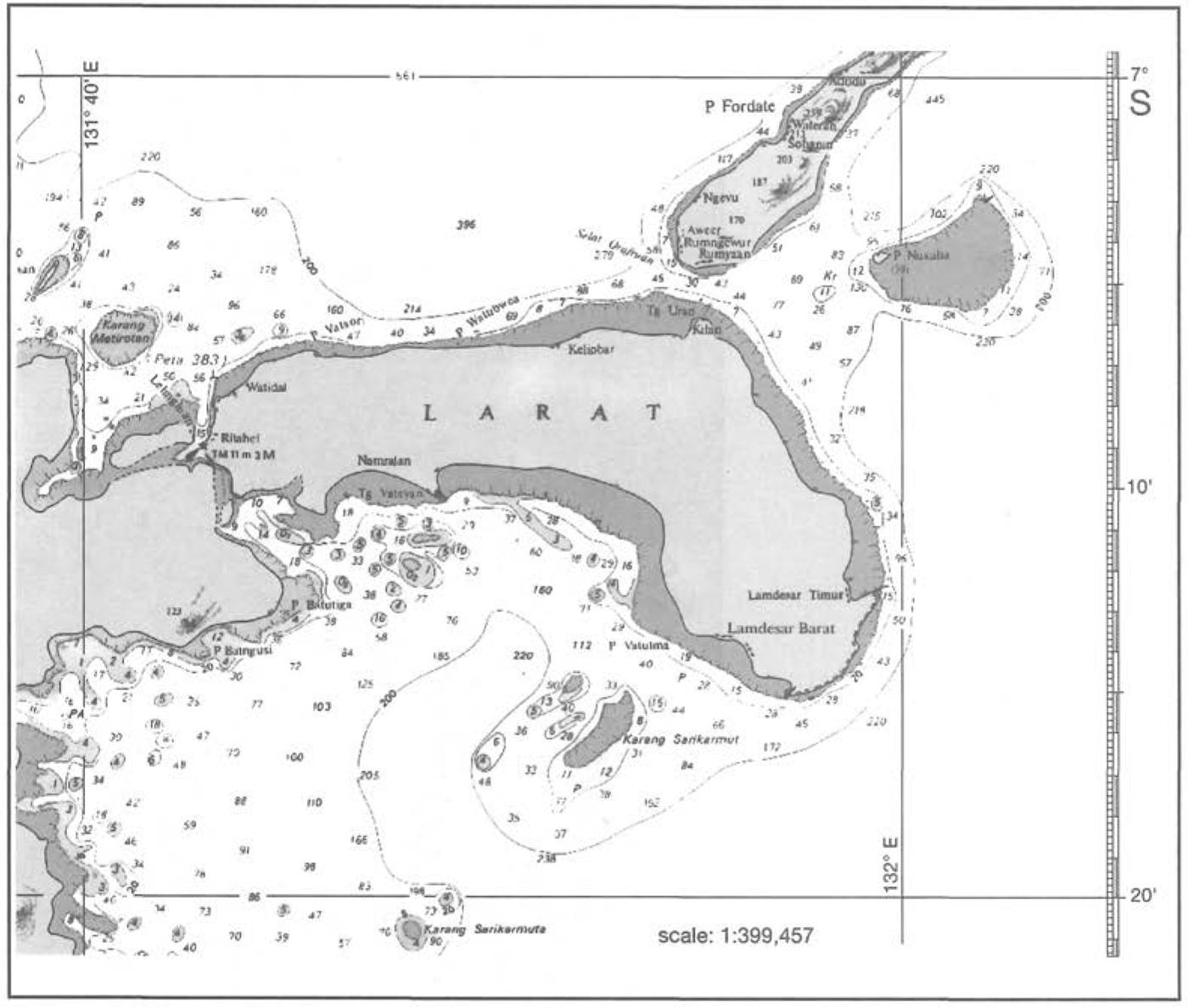
Figure 4.14. Location of Pulau Larat, Tanimbar Islands, illustrating the continuous nature of many fringing reefs in the archipelago. Note the large patch reef, Pulau Nukaha, to the northeast (upper right). This large patch reef (6.5 x 3.5 km) is often called an atoll, since it has a central lagoon that is not shown on the hydrographic chart. Note the small submerged barrier reef (c. 4 km long) on the south coast of P. Larat. Original scale 1:250,000.

Figure 4.15. A) Map of Bahuluang and Selayar Islands (upper right) from hydrographic chart No. 372. Soundings in metres. B) An oblique aerial photograph of Bahuluang Island (enclosed in A) showing that the fringing reef complex extends a considerable distance offshore (looking from north to south). Note the white sand cay at the northern limit of the reef complex.
Photo by Tomas and Anmarie Tomascik.
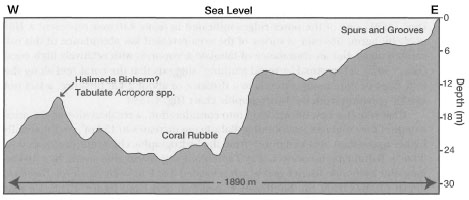
Figure 4.16. Depth profile of Bahuluang Island. Traced from an echo-sounding in van der Land and Sukarno (1987).
Bahuluang Island, forming the northern spur of the Tambolongang Platform, was visited by the Snellius-II Expedition, and, therefore, valuable information exists for an accurate areal estimate of the fringing reef. According to navigation chart No. 372 (scale 1:200,000), Pulau Bahuluang is surrounded by a fringing reef that is about 9 km in length and about 250 m in width (i.e., shore to reef crest). A peculiar 400-m-wide reef spur (or a spit, according to van der Land and Sukarno 1987) extends from the northeast tip of the island in a northerly direction for about 2 km. Using a simple calculation, it can be estimated that the fringing reef surrounding Bahuluang has an area of about 2 km2. Including the approximate 1 km2 area of the reef spur in the calculation will increase the total reef area of Bahuluang to about 3 km. The bathymetry of the area suggests that the west and east coast of the island have markedly different reef profiles. This has been confirmed by the Snellius-II Expedition scientists, who studied the zonation profile of the fringing reef on both coasts of the island. The fringing reef along the east coast is narrower (50100 m wide) when compared to the western coast, and is characterized by a sharp (45°-90°) drop-off at the seaward margin from a depth of about 4 m. According to van der Land and Sukarno (1987), the narrow reef is covered by large stands of branching Acropora spp., small foliaceous Montipora spp., as well as assemblages of massive Pontes spp. up to 1 m in diameter. Van Duyl (1991) reported large monospecific stands of Acropora microphthalma and Montipora foliosa at a depth of 4-18 m on a gentler slope at the southeast coast of the island, with coral cover exceeding 80%. In contrast, the profile of the west coast reef, illustrated in figure 4.16, is characterized by a gentler slope and a series of undulating terraces with distinct spurand-groove systems and extensive coral cover (van Duyl 1991).
Seaward of the reef slope are a series of coral ridges running parallel to the fringing reef, separated by a narrow channel (about 20 m in depth) with a coral rubble bottom. The walls of the channel are covered by corals. According to van Duyl (1991) one of the outer ridges indicated in figure 4.16 may represent a Halimeda bioherm; however, a survey of the area revealed low abundance of this calcareous algae, but an abundance of tabulate acroporids with relatively high coral cover (30%). Thus visual "ground truthing" suggests that the coral reef along the west coast of Bahuluang extends to a distance of about 2 km offshore, a fact not readily apparent from the hydrographic chart (fig. 4.15A).
If we take this new information into consideration, a recalculation of the areal extent of the coral reef surrounding Bahuluang increases to 15 km , a substantially higher figure than obtained purely from the hydrographic chart interpretation (i.e., 3 km2). Bahuluang is connected to Tambolongang Island, due south, by a 10-kmlong and 2-km-wide barrier reef which is located 3 to 9 m below sea level. Based on depth profiles from the Snellius-II Expedition and from figure 4.15B, we may assume that this 20 km2 area may also be classified as a coral reef system. It appears that the Tambolongang Platform, capping a southward trending submarine ridge, may be a large carbonate platform with an area of about 350 km. The eastern margin of the Tambolongang Platform is fringed by an almost continuous reef system that extends more than 30 km from the southern tip of Pulasi Island to the sand cay north of Bahuluang Island. The western margin of the Tambolongang Platform is either a partially drowned barrier reef, or a barrier reef in the early stages of development.
Another important factor that must be taken into consideration when evaluating the accuracy of the 7500 km2 coral reef area estimate is the length of the coastline used in the approximation. In fact, the length used was the straight baseline distance that defines Indonesian archipelagic boundaries (i.e., 80,791 km) (Moosa 1995a) and is, therefore, only loosely relevant to fringing reef area approximation. It has recently been suggested that the length of the Indonesian coastline, here defined as the line that forms the boundary between land and water and includes all 17,508 islands in the archipelago, may actually be as long as 204,000 km, second only to Canada, with a coastline of 243,797 km (Astuti et al. 1994). If the 204,000 km is used as the approximate length of the coastline and the same equation is used as for the 7500 km2 estimate, we arrive at a new rough estimate of 18,360 km2 for the areal extent of Indonesian fringing reefs.
However, it has recently been suggested that about 67% of the Indonesian coastline is fringed by coral reefs (Moosa 1995b), which is considerably higher than the original 45% figure. Taking this new figure at face value and further assuming that the fringing reefs are on an average only 200 m wide, two additional estimates are possible. If 80,791 km is used as the length of the archipelagic coastline then the area of fringing reefs is about 10,825 km2. However, if 204,000 km is used as the coastline length, then the area estimate of fringing reefs increases to 27,336 km2. This seemingly futile exercise clearly illustrates the need to determine the exact length of the coastline, the percentage of coastline fringed by a reef (i.e., fringing reef) and the average width of the coral reefs. Without these basic data the debate about the reefal area will continue to be based on pure guessing. Since other coral reefs, such as barrier reefs, patch reefs, platform reefs and atolls, are not included in either approximation, both values can be considered as serious underestimates.
To obtain an accurate figure for the Indonesian coral reef area coverage, there is a need for a clearer definition of a coral reef, since, as Crossland et al. (1991) state:
Estimation of the global [or regional] significance of coral reefs depends upon an ability to delineate them - spatially, temporally and functionally.
Clearly, the coral reef ecosystems do not consist of only intertidal reef flats. Extensive analysis of new hydrographic charts that are now available in various scales has allowed us to make a first rough approximation of coral reef area that includes barrier reefs, oceanic platform reefs and atolls. These data are summarized in tables 4.2, 4.3 and 4.4.
Using the new information, we may derive a new estimate for the areal approximation of coral reefs in the archipelago. To estimate the approximate area of fringing reefs in the archipelago we use the following equation:
FRA= [C(km)*0.45]*0.4(km)
Where 'C' is the length of the coastline (i.e., 80,791 km); '0.45' represents the 45% of coastline most likely covered by fringing reefs; and 0.4 km is the average width of the fringing reef. Thus, based on the above formula we arrive at a fringing reef areal estimate of about 14,542 km2. Table 4.5 provides the summary of the coral reef area approximations for the archipelago. Our conservative estimate for total coral reef area stands at roughly 85,700 km2, or about 14% of the world's total. Note that patch reefs associated with barrier reef lagoons are included within the barrier reef area estimate. However, offshore patch reef systems, such as the Kepulauan Seribu, for example, have not been included in the approximation.
Table 4.2. The number and approximate area of barrier reefs in the Indonesian Archipelago. West Indonesia: west coast of Sumatra, Java Sea, Malacca Strait, Natuna Sea, South China Sea, west Makassar Strait, west Sulawesi Sea. Central Indonesia: east Makassar Strait, east Sulawesi Sea, Flores Sea, Gulf of Tomini, west Molucca Sea, west Banda Sea, Flores Sea, Gulf of Bone, Indian Ocean (e.g., West Nusa Tenggara), Sawu Sea, Timor Sea. East Indonesia - Pacific Ocean, Arafura Sea, Banda Sea (east and central), Seram Sea, Halmahera Sea, Gulf of Irian (e.g., Cenderawasih Bay). Number of barrier reefs: total number of discrete barrier reefs per region. Lagoon width: average distance from shore to the seaward edge of the barrier reefs, based on maximum distances. Lagoon depth: average depth of the lagoon based on maximum depths. Length: total length of the barrier reefs. Based on hydrographic charts (Tentara Nasional Indonesia - Angkatan Laut, DINAS HIDRO - OSEANOGRAFI) with scales ranging from 1:250,000 to 1:15,000. Formula used for each barrier reef area approximation: Reefal Area (km2) = [Lagoon Width (km)/2] x [Length of the Barrier Reef (km)].
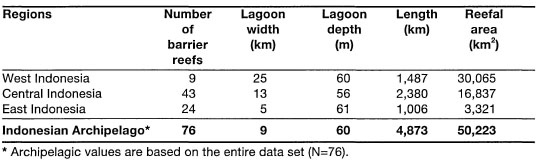
Table 4.3. Distribution of atolls in the Indonesian Archipelago. Reefal area approximations are based on the formula: Reefal Area (km2) = [Maximum Diameter (km)] x [Diameter Perpendicular to the Maximum Diameter (km)]. Geographic regions and methods as for table 4.2.

Table 4.4. Distribution of oceanic platform reefs (lagoonal and lagoonless) in the Indonesian Archipelago. Reefal area approximation based on the formula: Reefal Area (km2) = [Maximum Length of Reef (km)] x [Maximum Width of Reef (km)]. Note: Island area excluded from the reefal area approximation. Regions and methods as for table 4.2.
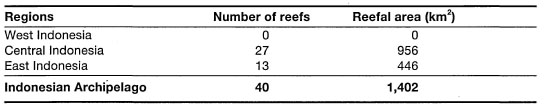
Table 4.5. An approximation of total reefal area in the Indonesian Archipelago associated with four main reef types (i.e., fringing reef, barrier reefs, platform reefs and atolls).